El ojo: III. Neurofisiologia central de la visión.

Vías visuales
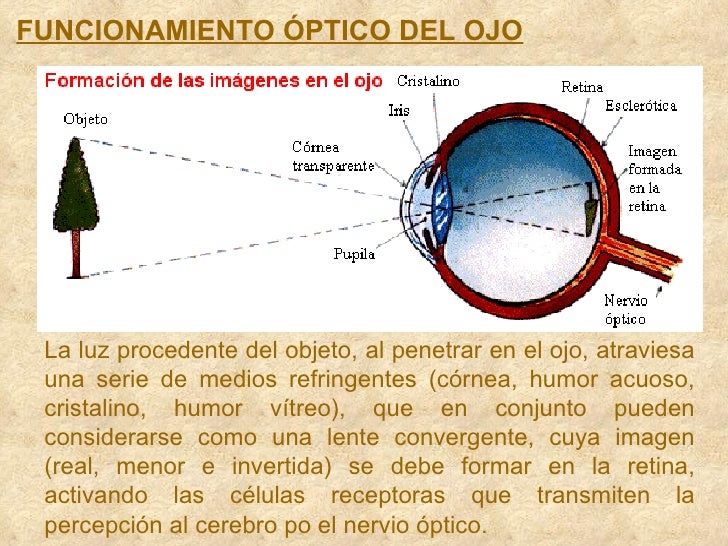
las principales vías visuales que van desde las dos retinas hasta la corteza visual. Las señales nerviosas de este carácter abandonan la retina a través de los nervios ópticos. En el quiasma óptico, las fibras procedentes de la mitad nasal de la retina cruzan hacia el lado opuesto, donde se unen a las fibras originadas en la retina temporal contraria para formar los tractos ópticos o cintillas ópticas. .as fibras de cada tracto óptico hacen sinapsis en el núcleo geniculado lateral dorsal del tálamo, y desde allí, las fibras geniculo calcar inas se dirigen a través de la radiación óptica (también denominada tracto geni- culo calcarino) hacia la corteza visual primaria en el área correspondiente a la cisura calcarina del lóbulo occipital medial. Las fibras visuales también se dirigen a otras regiones más antiguas del encéfalo: 1) desde los tractos ópticos llega hasta el núcleo supraquiasmático del hipotálamo, se supone que para regular los ritmos circadianos que sincronizan los diversos cambios fisiológicos del organismo según la noche y el día; 2) hacia los núcleos pretectales en el mesencèfalo, para suscitar movimientos reflejos de los ojos a fin de enfocarlos sobre los objetos de importancia y activar el reflejo fotomo- tor pupilar; 3) hacia el colículo superior, para controlar los movimientos direccionales rápidos de ambos ojos, y 4) hacia el núcleo geniculado lateral ventral del tálamo y las regiones basales adyacentes del cerebro, se cree que para contribuir al dominio de algunas de las funciones conductuales que lleva a cabo el organismo. Así pues, las vías visuales pueden dividirse a grandes rasgos en un sistema antiguo dirigido hacia el mesencèfalo y la base del prosencéfalo y un sistema nuevo para la transmisión directa de las señales visuales hacia la corteza visual situada en los lóbulos occipitales

Función del núcleo geniculado lateral dorsal del tálamo
Ese núcleo es una estructura laminar que consta de seis capas concéntricas La más interna es la capaI, y la más superficial,la capa VI. Los axones de la retina que terminan en este núcleo provienen de la retina nasal contralateral y de la retina temporal ipsolateral, y transportan, en consecuencia, una información punto a punto del campo visual contralateral. Las fibras nasales contralaterales terminan en las capas I, IV y VI, y las fibras temporales ipsolaterales, en las capas II, III y V. La información de ambos ojos sigue segregada en el núcleo geniculado lateral dorsal, al igual que las señales provenientes de las células ganglionares X e Y de la retina La entrada de las células Y termina en las capas I y II, que se denominan capas magno celulares porque contienen neuronas bastante grandes. Esta es una vía de conducción rápida, con ceguera para los colores, pero con una información localizadora eficaz. Las capas III a VI se denominan parvo celulares porque contienen neuronas bastante pequeñas que reciben señales de las células X, que transmiten información del color y de la forma. Por tanto, la información de la retina se procesa a lo largo de al menos dos vías paralelas: 1) una corriente dorsal que transporta información de los bastones y las grandes células ganglionares(Y)que dan información específica de la localización y del movimiento, y 2) una corriente ventral que transporta información del color y de la forma desde los conos y las pequeñas células ganglionares (X).

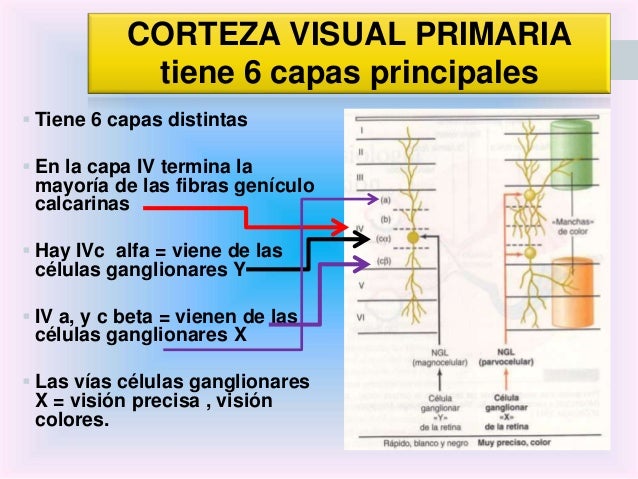
Organización y función de la corteza visual
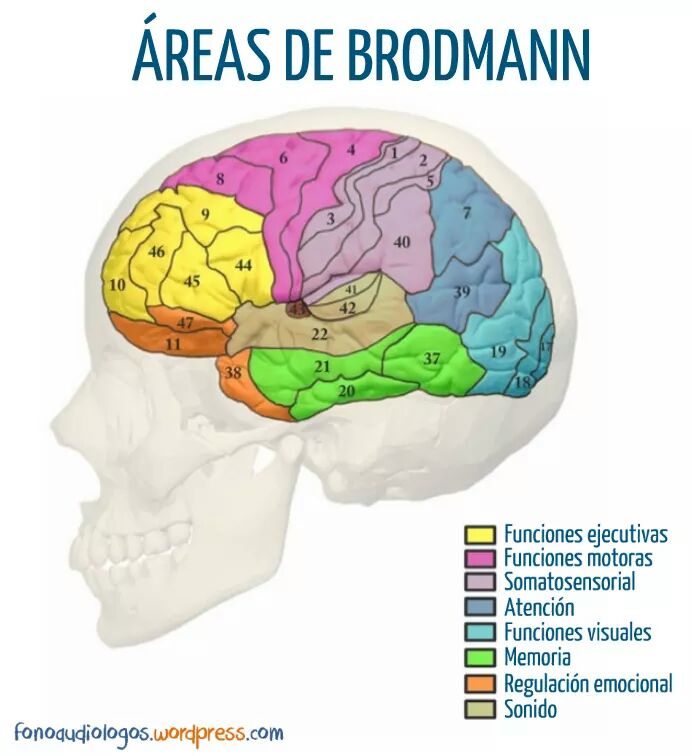
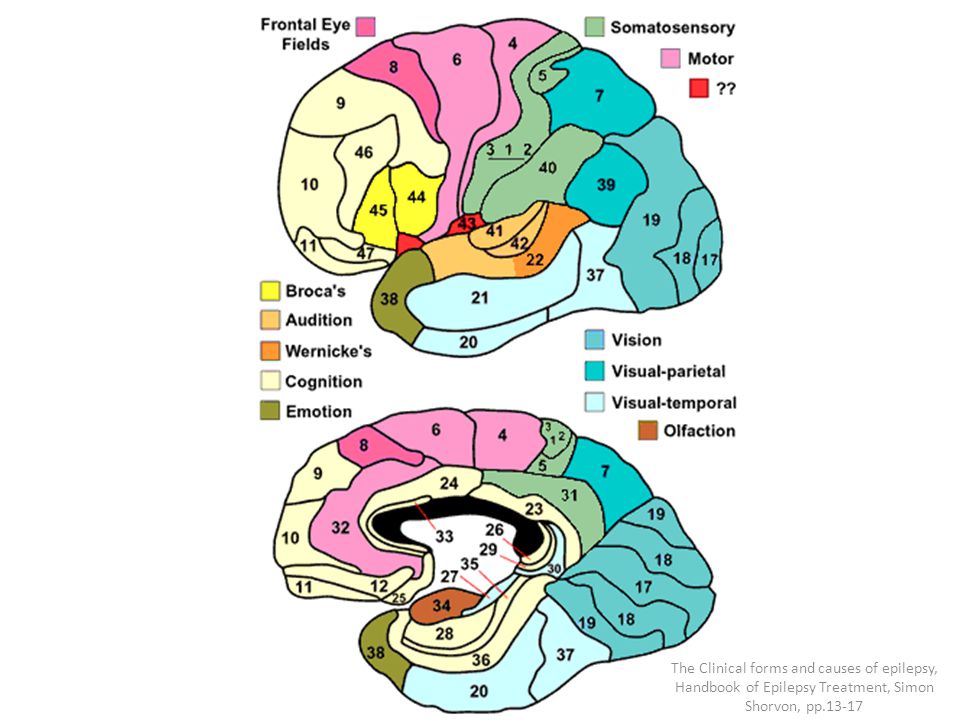
La corteza visual primaria, o área 17 de Brodmann ,también se denomina V-1.Está localizada en la cara medial del hemisferio y reviste las dos paredes del surco calcarino, cerca del polo occipital. Recibe señales visuales de cada ojo y contiene la representación de todo el campo visual contralateral: el campo visual inferior está contenido en el banco superior del surco calcarino,y el superior,en el inferior. La porción macular de la retina se representa posteriormente,cerca del polo occipital,y las señales de la retina más periférica alcanzan los territorios más anteriores. La corteza visual secundaria (denominada V-2 a V-5) rodea el área primaria y se corresponde con las áreas 18 y 19 de Brodmann, con la circunvolución temporal media (TM) y con las áreas 7a y 37 de Brodmann. La corteza visual primaria se estructura en capas. Como todas las demás áreas de la neo corteza, la corteza visual primaria se organiza en seis capas horizontales. Las fibras geniculadas de entrada, de tipo Y, terminan, principalmente, en una subdivisión de la capa IV denominada IVca ,mientras que las de tipo X terminan principalmente en las capas IVa y IVcb

Columnas neuronales verticales en la corteza visual.
La corteza visual posee una organización estructural formada por varios millones de columnas verticales de células neuronales, con un diámetro de 30 a 50 |xm cada una. Esta misma disposición columnar vertical se observa por toda la corteza cerebral dedicada a las demás modalidades sensitivas (y también por las regiones corticales motoras y de análisis). Cada columna constituye una unidad funcional. A grandes rasgos puede calcularse que una columna vertical visual tal vez contenga 1.000 neuronas o más. Una vez que las señales ópticas llegan a la capa IV, sufren una nueva transformación al propagarse hacia el exterior y hacia el interior a lo largo de cada unidad columnar vertical. Se cree que este procesamiento descifra componentes independientes de la información visual en las estaciones sucesivas a lo largo de la vía. Los impulsos que ascienden hacia las capas I, II y III acaban enviando su contenido en sentido lateral con un alcance reducido dentro de la corteza. Por el contrario, los que descienden hacia las capas V y VI excitan neuronas que transmiten su actividad a unas distancias mucho mayores.
«Manchas de color» en la corteza visual.
Intercaladas entre las columnas visuales primarias y también entre las columnas de algunas áreas visuales secundarias hay unas regiones especiales de tipo columnar llamadas manchas de color; estas zonas reciben señales laterales desde las columnas visuales adyacentes y se activan de forma específica por los estímulos de color. Por tanto, se supone que constituyen las zonas primarias para descifrar el color.
Interacción de las señales visuales procedentes de ambos ojos.
Recuerde que las señales visuales procedentes de los dos ojos se transmiten a través de capas neuronales independientes en el núcleo geniculado lateral. Estos impulsos aún permanecen separados entre sí cuando llegan a la capa IV de la corteza visual primaria. En realidad, la capa IV está entrelazada con las bandas de las columnas neuronales, cuya anchura mide unos 0,5 mm; la actividad procedente de un ojo penetra en las columnas de una banda sí y otra no, según una disposición alterna con las señales del segundo ojo. Este área cortical descifra si las zonas respectivas de las imágenes visuales originadas en cada ojo están en concordancia entre sí es decir, si los puntos correspondientes de las dos retinas coinciden entre ellos). A su vez, la información descifrada se emplea para corregir la mirada direccional de ambos ojos con el fin de que se fusionen entre sí (se pongan en concordancia). Los datos observados sobre el grado de coincidencia entre las imágenes de ambos ojos también permiten que una persona detecte la distancia a un objeto por el mecanismo de la estereopsia.


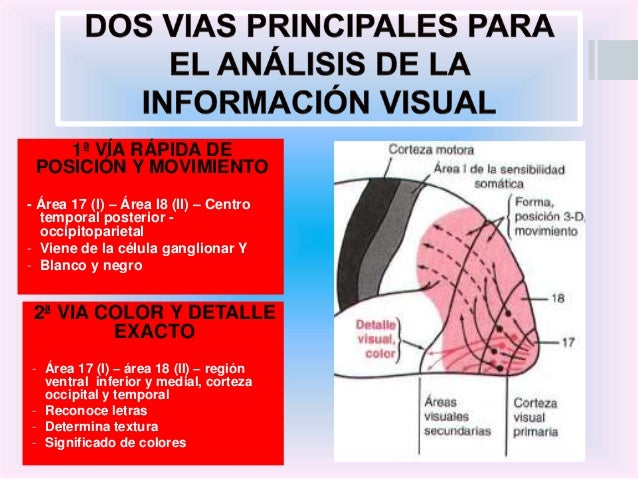
Dos vías importantes para el análisis de la información visual: 1) la vía rápida de la «posición» y el «movimiento», y 2) la vía de la exactitud del color
1.Análisis de la posición tridimensional, la forma global y el movimiento de los objetos. Una de las vías de análisis, examina la posición tridimensional que ocupan los objetos visuales en el espacio que rodea al organismo. Esta vía también explora la forma física global de la escena visual, así como el movimiento que se produce en su seno. Dicho de otro modo, dice dónde está cada objeto en cada instante y si está en movimiento o no. Después de salir de la corteza visual primaria, los impulsos viajan en general hacia el área temporal media posterior y ascienden hacia la extensa corteza occipitoparietal. En el borde anterior de la corteza parietal se superponen con los que proceden de las áreas de asociación somáticas posteriores dedicadas a analizar los aspectos tridimensionales de las señales somatosensitivas. La información contenida en esta vía encargada de la posición-forma-movimiento básicamente viene de las fibras Y grandes del nervio óptico originadas en las células ganglionares Y de la retina, que transmiten impulsos rápidos pero únicamente de carácter descriptivo en blanco y negro sin color.
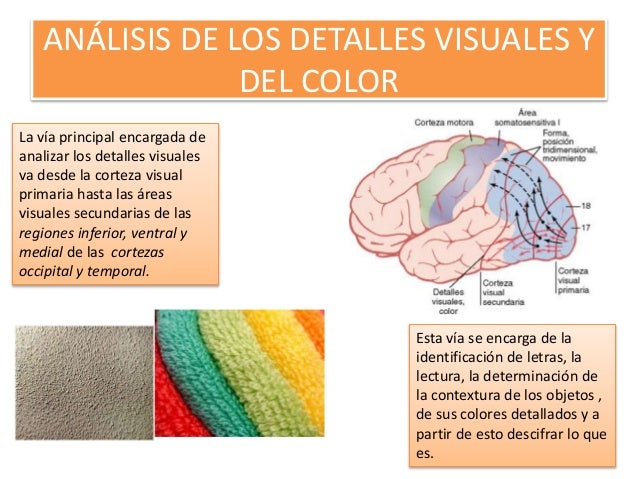
2.Análisis de los detalles visuales y del color que van desde la corteza visual primaria hasta las áreas visuales secundarias de las regiones inferior, ventral y medial de las cortezas occipital y temporal, muestran la vía principal encargada de analizar los detalles visuales. Otras zonas aparte de esta misma vía también se dedican a analizar específicamente el color. Por tanto, esta vía se ocupa de proezas visuales como la identificación de las letras, la lectura, la determinación de la textura de los objetos, de sus colores detallados, y de descifrar lo que es y lo que significa un objeto a partir de toda esta información.

Patrones neuronales de estimulación durante el análisis de una imagen visual
La corteza visual también detecta la orientación de las líneas y los bordes: las células simples. La corteza visual no sólo detecta la existencia de líneas y bordes en las diferentes zonas de la imagen retiniana, sino también la dirección en la que se están orientadas cada una de ellas, es decir, si son verticales u horizontales o si guardan un cierto grado de inclinación. Se cree que esto obedece a la excitación de neuronas de segundo orden por parte de sistemas lineales de células que se inhiben mutuamente, cuando esta inhibición se produce siguiendo toda la línea celular que corresponde a un límite de contraste. Por tanto, con cada orientación de una línea de este tipo, se estimulan células neuronales específicas. Una línea dispuesta según una dirección diferente excita un grupo de células distinto. Estas neuronas se denominan células simples. Sobre todo están situadas en la capa IV de la corteza visual primaria
Detección de la orientación lineal cuando una línea se desplaza en sentido lateral o vertical a lo largo del campo visual: las células «complejas».
A medida que la señal visual se aleja de la capa IV, algunas neuronas responden a líneas orientadas en la misma dirección pero sin ser específicas de una posición. Esto es, incluso si una línea recorre por el campo una distancia moderada en sentido lateral o vertical, estas mismas neuronas poco abundantes aún seguirán estimuladas si la línea conserva la misma dirección. Dichas neuronas se llaman células complejas.
Detección de líneas con una longitud, un ángulo u otras formas específicas.
Algunas neuronas de las capas más externas de las columnas visuales primarias, lo mismo que las pertenecientes a ciertas áreas visuales secundarias, no se estimulan más que por líneas o bordes con una longitud o una forma en ángulo específicas, o por imágenes que poseen otras características. Es decir, estas neuronas detectan órdenes aún más elevadas de información procedentes de la escena visual. Por tanto, a medida que uno asciende dentro de la vía analítica
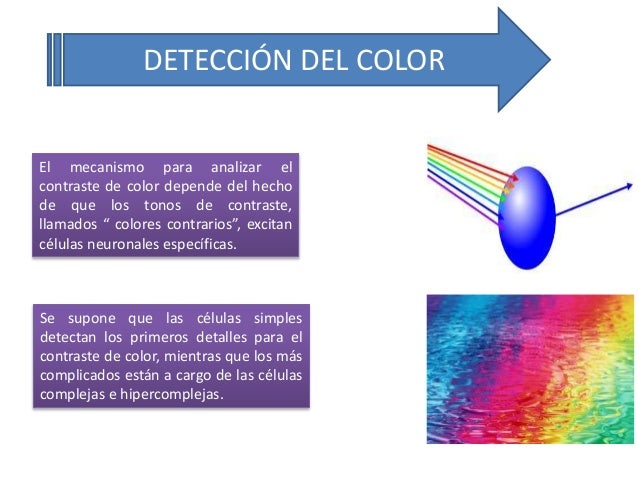
Detección del color
El color se detecta básicamente por el mismo procedimiento que las líneas: mediante su contraste. Por ejemplo, una zona roja suele quedar contrastada frente a otra verde, una azul contra una roja, o una verde contra una amarilla. Todos estos colores también pueden compararse frente a una zona blanca en el seno de la escena visual. De hecho, se piensa que este contraste con el blanco es el responsable principal del fenómeno llamado constancia del color; a saber, cuando el color de un haz de iluminación cambia, también lo hace el tono del blanco con esta luz, y el cálculo oportuno por parte del cerebro permite interpretar el rojo como rojo aun cuando el haz de iluminación haya modificado el color que llega a los ojos. El mecanismo para analizar el contraste de color depende del hecho de que los tonos de contraste, llamados colores contrarios, excitan células neuronales específicas. Se supone que las células simples detectan los primeros detalles para el contraste de color, mientras que los más complicados están a cargo de las células complejas e hipercomplejas.

Campos visuales; campimetría
El campo visual es la zona de visión observada por un ojo en un instante dado. La región percibida por el lado nasal se llama campo visual nasal y la que llega al lado lateral campo visual temporal. Para diagnosticar una ceguera en una porción específica de la retina, se cartografía el campo visual de cada ojo mediante un procedimiento llamado campimetría. Esto se consigue haciendo que el individuo mire con un ojo cerrado y dirija el otro hacia un punto central situado directamente delante de él. A continuación, se desplaza un foco de luz o un objeto de pequeñas dimensiones hacia adelante y hacia atrás por todas las zonas del campo visual, y la persona indica cuándo puede verlo y cuándo no. De esta manera, se traza el campo visual del ojo izquierdo . En todos los trazados campimétri- cos aparece un punto ciego ocasionado por la ausencia de conos y bastones en la retina sobre el disco óptico que queda a unos 15° lateral al punto central de la visión


Efecto de las lesiones de la vía óptica sobre los campos visuales.
La destrucción de todo el nervio óptico origina una ceguera en el ojo afectado. Si el trastorno afecta al quiasma óptico impide el cruce de los impulsos que proceden de la mitad nasal de cada retina y van dirigidos hacia el tracto óptico del lado opuesto. Por tanto, esta mitad queda ciega a ambos lados, lo que significa que la persona está ciega en el campo temporal de cada ojo porque la imagen del campo visual se encuentra invertida en la retina debido al sistema óptico ocular; esta situación se denomina hemianopsia bitemporal. Tales lesiones normalmente son el resultado de un tumor hipofisario que presiona sobre la parte inferior del quiasma desde la silla turca hacia arriba. La interrupción de un tracto óptico deja sin inervación la mitad de cada retina correspondiente al mismo lado de la lesión; como consecuencia, ningún ojo es capaz de ver los objetos situados en el lado opuesto de la cabeza. Dicha situación se conoce como hemianopsia homónima.



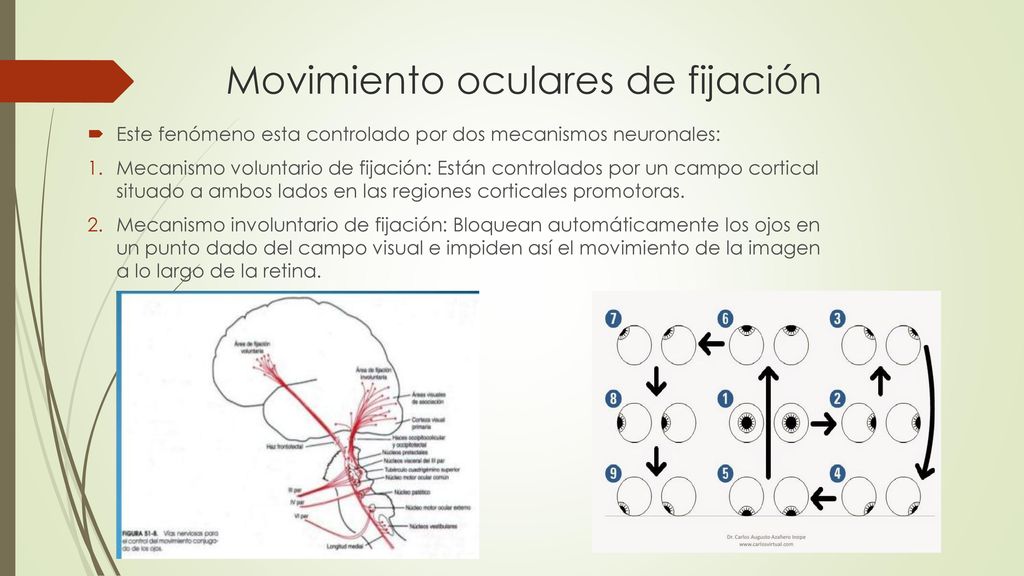
Movimientos oculares de fijación
Tal vez los movimientos más importantes de los ojos son los que provocan su fijación en una porción concreta del campo visual. Este fenómeno está controlado por dos mecanismos neuronales. El primero permite a una persona mover los ojos voluntariamente para encontrar el objeto sobre el que desea fijar la visión; esto se llama mecanismo voluntario de fijación. El segundo es un proceso involuntario que mantiene los ojos fijos con firmeza sobre el objeto una vez que ha sido descubierto; esto se denomina mecanismo involuntario de fijación
Los movimientos voluntarios de fijación están controlados por un campo cortical situado a ambos lados en las regiones corticales premotoras, La disfunción o destrucción bilateral de estas áreas complica ia posibilidad de que una persona desbloquee su mirada de un punto de fijación y la cambie hacia otro o vuelve esta maniobra casi imposible. Suele ser necesario parpadear o taparse los ojos con una mano un breve instante, lo que ya sí que permite mover los ojos. Por el contrario, el mecanismo de fijación que provoca el «bloqueo» de los ojos sobre el objeto de atención una vez que se ha descubierto su presencia está controlado por las áreas visuales secundarias de la corteza occipital, situadas básicamente por delante de la corteza visual primaria. Cuando en un animal se provoca una destrucción bilateral de este área de fijación, esta circunstancia le crea problemas para mantener los ojos dirigidos hacia un punto de fijación dado o puede incapacitarle por completo para hacerlo. En resumen, los campos oculares involuntarios en la zona posterior de la corteza occipital bloquean automáticamente los ojos en un punto dado del campo visual e impiden así el movimiento de la imagen a lo largo de la retina. Para desbloquear esta fijación visual han de transmitirse señales voluntarias desde los campos oculares corticales voluntarios situados en las cortezas frontales.
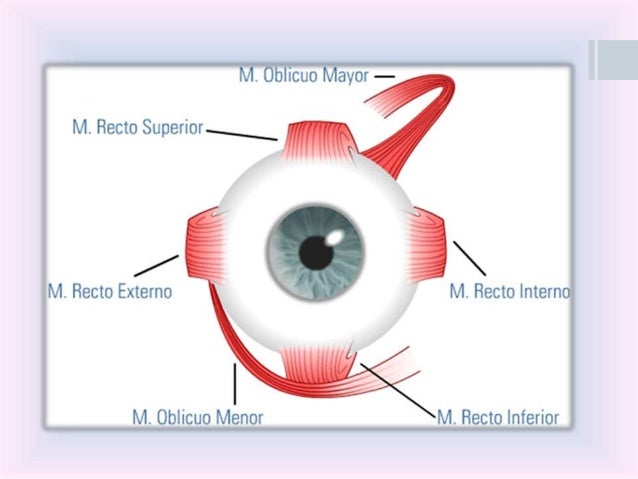
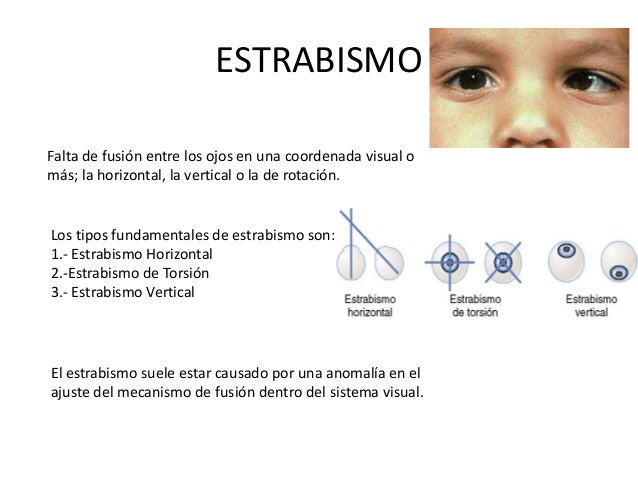
«Fusión» de las imágenes visuales de ambos ojos
Para que las percepciones visuales resulten más elocuentes, normalmente se fusionan entre sí las imágenes visuales de ambos ojos según los «puntos correspondientes» de las dos retinas. La corteza visual ocupa un lugar importante en el proceso de fusión. Ya se indicó antes en este mismo capítulo que los puntos correspondientes de ambas retinas transmiten señales visuales hacia capas neuronales diferentes en el cuerpo geniculado lateral y a su vez estos impulsos se transfieren hasta las neuronas situadas en una posición paralela en la corteza visual. Se producen interacciones entre estas neuronas corticales que dan lugar a la excitación por interferencia de neuronas específicas cuando las dos imágenes visuales no quedan en concordancia, es decir, cuando su fusión no se realiza con precisión. Se supone que esta excitación suministra la señal que se transmite al aparato oculomotor para provocar la convergencia y la divergencia o la rotación de los ojos a fin de que pueda restablecerse la fusión. Una vez que coinciden los puntos correspondientes de las dos retinas, desaparece la excitación de las neuronas específicas de interferencia en la corteza visual

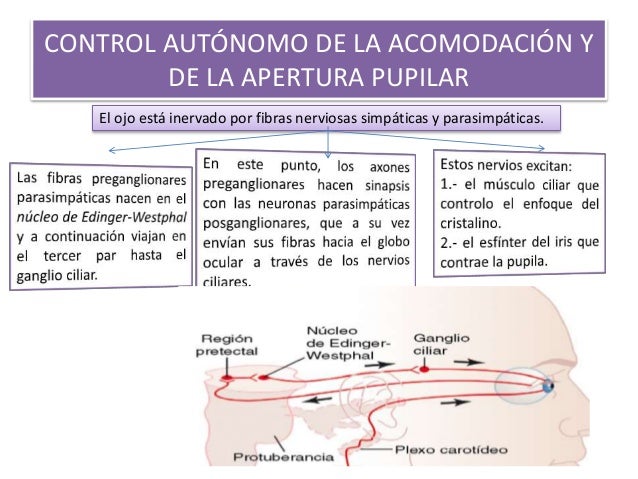



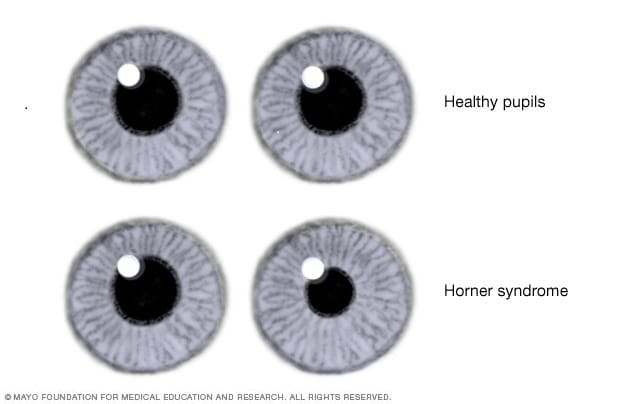
Síndrome de Horner
Los nervios simpáticos del ojo a veces quedan interrumpidos. Esta circunstancia suele suceder en la cadena simpática cervical. Esto provoca el cuadro clínico llamado síndrome de Horner, que ocasiona los siguientes efectos. Primero, debido a la interrupción de las fibras nerviosas simpáticas dirigidas al músculo dilatador de la pupila, esta permanece contraída de forma continua con un diámetro más pequeño que la pupila del lado opuesto. Segundo, el párpado superior se cae debido a que normalmente se mantiene en posición abierta durante las horas de vigilia en parte por la contracción de las fibras musculares lisas contenidas en su interior e inervadas por el sistema simpático. Por tanto, la destrucción de estos nervios imposibilita su apertura hasta una altura normal. Tercero, los vasos sanguíneos del lado correspondiente de la cara y de la cabeza quedan dilatados de un modo persistente. Cuarto, no puede producirse la sudoración (que requiere la acción de las señales nerviosas simpáticas) en el mismo lado de la cara y de la cabeza afectado por el síndrome de Horner







/231053764009/231053764009_gf30.png)