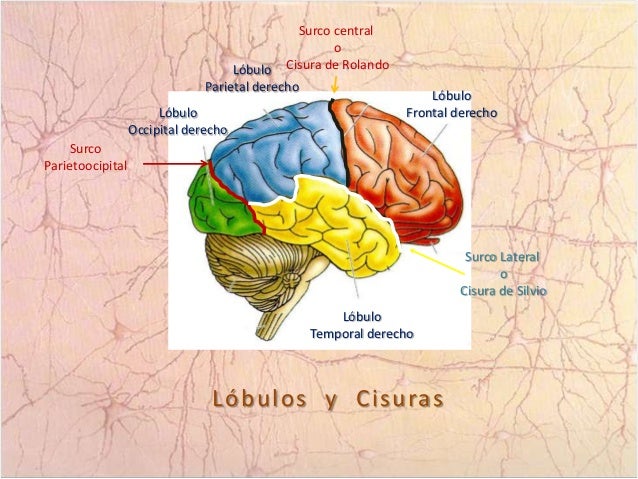
Flujo sanguíneo cerebral.
El flujo sanguíneo en el encéfalo es suministrado por cuatro grandes arterias, dos carotídeas y dos vertebrales, que se funden para formar el polígono de Willis en la base del encéfalo. Las arterias que parten del polígono de Willis se desplazan a lo largo de la superficie cerebral y dan origen a las arterias piales, que se ramifican en vasos más pequeños denominados arterias y arteriolas penetrantes.
Los vasos penetrantes están separados ligeramente del tejido encefálico por una extensión del espacio subaracnoideo denominada espacio de Virchow-Robin. Los vasos penetrantes se sumergen en el tejido encefálico, para dar lugar a arteriolas intracerebrales, que a su vez se ramifican en capilares en los que tiene lugar el intercambio entre la sangre y los tejidos de oxígeno, nutrientes, dióxido de carbono y metabolitos.
Regulación del flujo sanguíneo cerebral.
Por término medio, el flujo sanguíneo normal a través del cerebro de una persona adulta es de 50 a 65 ml cada 100 g de tejido por minuto. Para todo el encéfalo, esta cantidad asciende 750 a 900 ml/min. Así pues, el encéfalo constituye únicamente en torno al 2% del peso corporal, pero recibe el 15% del gasto cardíaco en reposo. Al igual que sucede en la mayoría del resto de tejidos, el flujo sanguíneo cerebral está muy relacionado con el metabolismo tisular.
Según se cree, varios factores metabólicos contribuyen a la regulación del flujo sanguíneo cerebral: 1) la concentración de dióxido de carbono; 2) la concentración de iones hidrógeno, 3) la concentración de oxígeno, y 4) sustancias liberadas de los astrocitos, que son células no neuronales especializadas que parecen acompañar la actividad neuronal con la regulación del flujo sanguíneo local.
Importancia del control ejercido por el dióxido de carbono y los iones hidrógeno sobre el flujo sanguíneo cerebral.
Una concentración alta de iones hidrógeno reduce mucho la actividad neuronal. Por tanto, es una suerte que su incremento también provoque un aumento del flujo sanguíneo, que a su vez retira del tejido cerebral iones hidrógeno, dióxido de carbono y otras sustancias formadoras de ácidos. La pérdida de dióxido de carbono elimina ácido carbónico de los tejidos; esta acción, junto con la extracción de otros ácidos, normaliza la concentración de iones hidrógeno. Por tanto, dicho mecanismo sirve para mantener una concentración constante de iones hidrógeno en los líquidos cerebrales y ayuda así a conservar la actividad neuronal a un nivel normal y constante.

La falta de oxígeno como factor regulador del flujo sanguíneo cerebral
Los experimentos han demostrado que el descenso en la presión parcial de oxígeno (Po2) del tejido cerebral por debajo de unos 30 mmHg (el valor normal es de 35 a 40 mmHg) comienza de inmediato a incrementar el flujo sanguíneo que recibe. Esto no deja de ser una suerte, pues el funcionamiento cerebral sufre una perturbación a valores no mucho menores de la Po2, especialmente si llega a menos de 20 mmHg. A estos niveles tan bajos puede aparecer incluso un coma. Por tanto, el mecanismo de regulación local sobre el flujo sanguíneo cerebral por parte del oxígeno constituye una respuesta protectora muy importante contra el descenso de la actividad neuronal cerebral y, en consecuencia, contra cualquier trastorno en la capacidad mental.
Medición del flujo sanguíneo cerebral y sus efectos sobre la actividad cerebral.
Se ha concebido un método para registrar el flujo sanguíneo en un mínimo de 256 segmentos aislados de la corteza cerebral humana al mismo tiempo. Para registrar el flujo sanguíneo en estos segmentos se inyecta en la arteria carótida una sustancia radiactiva, como xenón radiactivo; a continuación, se recoge la radiactividad de cada segmento cortical a medida que la sustancia atraviesa el tejido cerebral. Con este fin, se ajustan 256 pequeños detectores de radiación contra la superficie de la corteza. La rapidez del ascenso y declive de la radiactividad en cada segmento tisular aporta una medida directa de la velocidad del flujo sanguíneo que lo atraviesa.

Microcirculación cerebral.
Una característica estructural importante que presentan los capilares del encéfalo es que en su mayoría son menos «permeables» que los capilares sanguíneos casi de cualquier otro tejido del organismo. Una razón para este fenómeno radica en que cualquiera de sus caras se encuentra reforzada por los «podocitos neurogliales», que consisten en pequeñas prolongaciones procedentes de las células de la glía a su alrededor, que lindan con todas las superficies de los capilares y suministran un soporte físico para impedir su estiramiento excesivo en el caso de que suba demasiado la presión sanguínea capilar.
Las paredes de las arteriolas pequeñas que conducen hacia los capilares del encéfalo acaban muy engrosadas en las personas que sufren una elevación de la presión sanguínea, y permanecen notablemente contraídas todo el tiempo para impedir que esta situación se transmita a los capilares. Más adelante en este mismo capítulo veremos que siempre que fracasan estos sistemas protectores contra la trasudación de líquido hacia el encéfalo, sobreviene un edema cerebral grave, que puede llevar con rapidez al coma y a la muerte.
El «ictus» cerebral aparece cuando se obstruyen los vasos sanguíneos cerebrales.
Casi todos los ancianos tienen bloqueadas algunas arterias pequeñas del encéfalo y hasta el 10% acaba sufriendo a la larga un bloqueo suficiente como para ocasionar un trastorno grave del funcionamiento cerebral, proceso llamado «ictus». La mayoría de los ictus están causados por placas arterioescleróticas que aparecen en una o más de las arterias que irrigan el encéfalo. Las placas tienen la capacidad de activar el mecanismo de la coagulación sanguínea, haciendo que se forme un coágulo y se bloquee el flujo sanguíneo en la arteria, lo que lleva a la pérdida súbita de las funciones cerebrales en un área circunscrita.

Más o menos en la cuarta parte de las personas que sufren un ictus, la presión arterial elevada hace que se rompa uno de los vasos sanguíneos; a continuación, se produce una hemorragia, que comprime el tejido cerebral local y altera así su funcionamiento. Los efectos neurológicos de un ictus vienen determinados por la zona afectada del encéfalo. Uno de los tipos más frecuentes es el bloqueo de la arteria cerebral media que irriga la porción intermedia de un hemisferio cerebral.
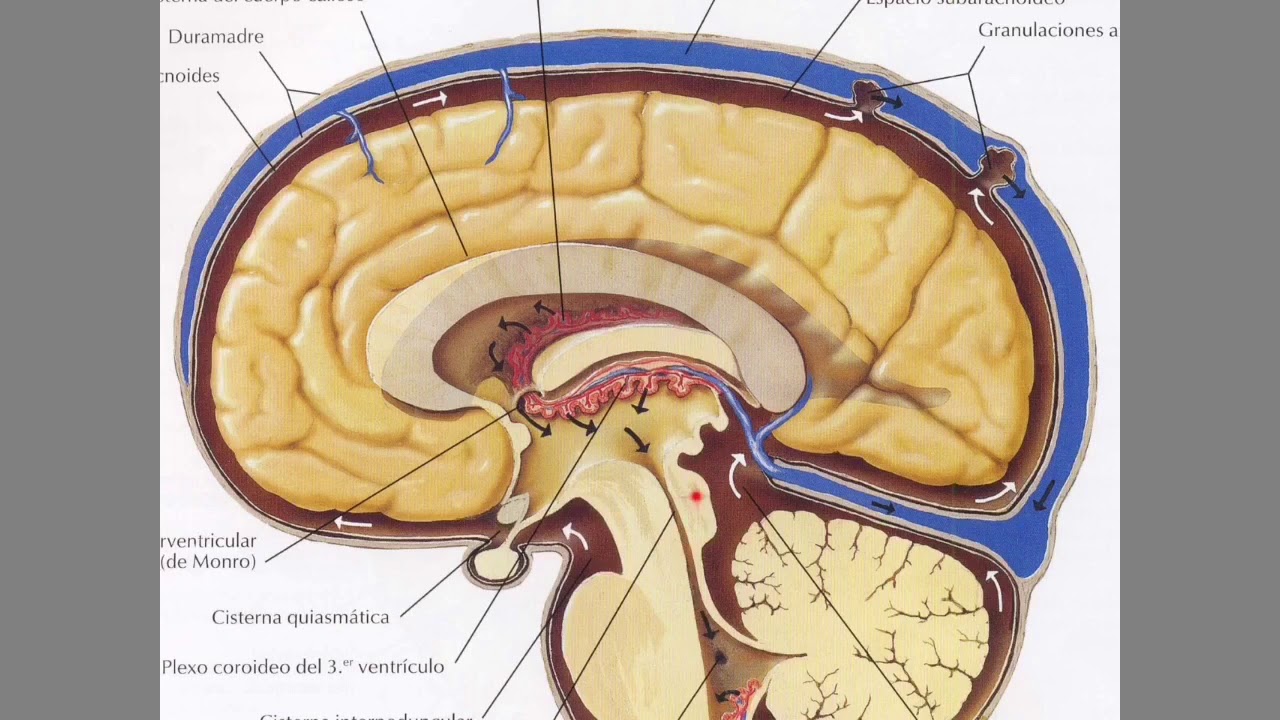
Sistema del líquido cefalorraquídeo.
Toda la cavidad que encierra el encéfalo y la médula espinal tiene una capacidad de unos 1.600 a 1.700 ml. De ellos, más o menos 150 ml están ocupados por el líquido cefalorraquídeo, y el resto por el encéfalo y la médula. Este líquido está presente en los ventrículos cerebrales, en las cisternas que rodean por fuera al encéfalo y en el espacio subaracnoideo alrededor del encéfalo y de la médula espinal. Todas estas cavidades se encuentran conectadas entre sí y la presión del líquido se mantiene a un nivel sorprendentemente constante.

Función amortiguadora del líquido cefalorraquídeo.
Una función fundamental del líquido cefalorraquídeo consiste en amortiguar el encéfalo dentro de su bóveda sólida. El encéfalo y el líquido cefalorraquídeo poseen aproximadamente la misma densidad específica (tan solo difieren en un 4% más o menos), de modo que el encéfalo se limita a flotar en el seno del líquido. Por tanto, un golpe en la cabeza, si no es demasiado fuerte, desplaza todo el encéfalo a la vez que el cráneo, lo que evita que cualquier porción suya sufra una torsión transitoria por su acción.
Formación, flujo y absorción del líquido cefalorraquídeo.
El líquido cefalorraquídeo se forma a una velocidad de unos 500 ml diarios, lo que supone el triple o el cuádruple de su volumen total en todo el sistema. Alrededor de dos tercios o más de esta cantidad se debe a la secreción desde los plexos coroideos en los cuatro ventrículos, sobre todo en los dos ventrículos laterales. Un poco más se produce en la superficie ependimaria de todos los ventrículos y en la aracnoides. Un pequeño porcentaje procede del encéfalo a través de los espacios perivasculares que quedan alrededor de los vasos sanguíneos que atraviesan el encéfalo.

La parte segregada en los ventrículos laterales pasa primero hacia el tercer ventrículo; después, tras la incorporación de una mínima cantidad más en esta cavidad, desciende a lo largo del acueducto de Silvio hacia el cuarto ventrículo, donde aún se añade otra minúscula proporción de líquido. Finalmente, sale del cuarto ventrículo por tres pequeños orificios, los dos agujeros laterales de Luschka y el agujero central de Magendie, para penetrar en la cisterna magna, un espacio de líquido que queda detrás del bulbo raquídeo y debajo del cerebelo. La cisterna magna se continúa con el espacio subaracnoideo que rodea al encéfalo y la médula espinal en su integridad. Casi todo el líquido cefalorraquídeo asciende a continuación desde la cisterna magna a través de estos espacios subaracnoideos alrededor del cerebro. Desde aquí, penetra por las múltiples vellosidades aracnoideas que sobresalen hacia el gran seno venoso sagital y otros senos venosos cerebrales, y las atraviesa. Por tanto, todo el líquido sobrante se vierte hacia la sangre venosa a través de los poros de estas vellosidades.
Metabolismo cerebral.
En condiciones de vigilia en reposo, al metabolismo cerebral le corresponde aproximadamente el 15% del metabolismo total del organismo, aunque su masa no supone más que el 2% de la masa corporal íntegra. Por tanto, en condiciones de reposo, el metabolismo cerebral por unidad de masa tisular es unas 7,5 veces el metabolismo medio que existe fuera de los tejidos del sistema nervioso.
El encéfalo no es capaz de efectuar un gran metabolismo anaerobio. Una de las razones para ello estriba en el elevado índice metabólico de las neuronas, por lo que la mayor parte de la actividad neuronal depende de la liberación de oxígeno cada segundo desde la sangre. Si se reúnen todos estos factores, puede entenderse por qué la interrupción brusca del flujo sanguíneo hacia el encéfalo o la ausencia total súbita de oxígeno en la sangre pueden provocar la pérdida del conocimiento en un plazo de 5 a 10 s.






















/Anim_Alzheimers-b64b3759b8334d30b2fd76cdd5d39d7f.gif)