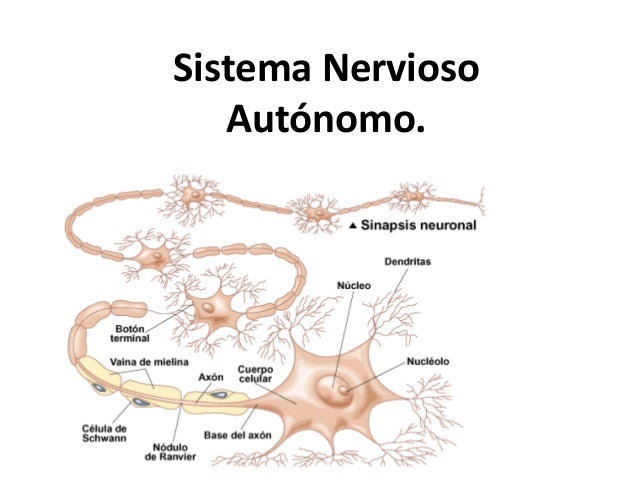
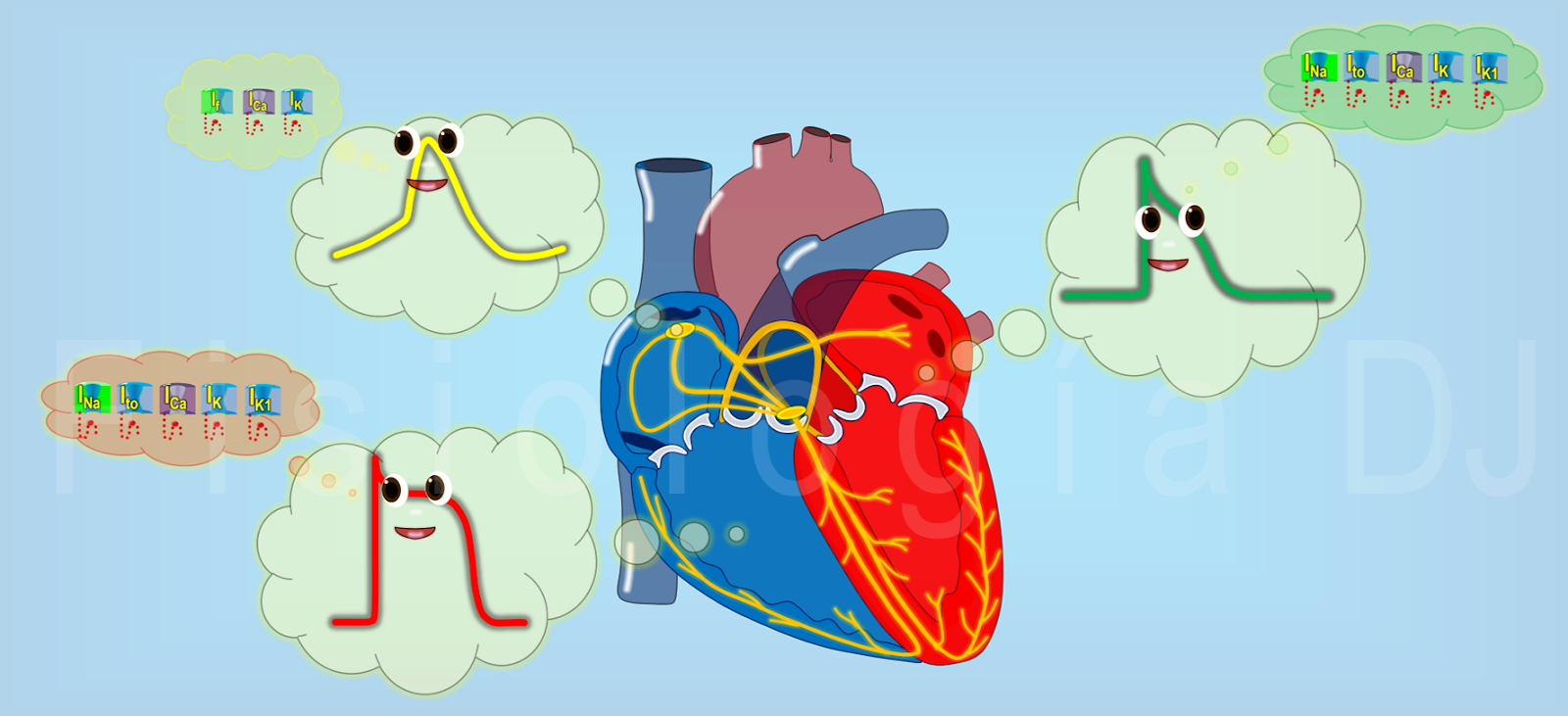
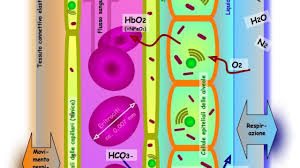
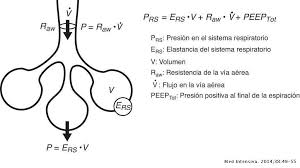
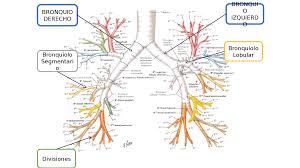
Sistema electrogenico del corazón: Generar impulsos rítmicos para producir contracciones rítmicas y conducir impulsos con rapidez. Sistema cardionector está formado por: Nodo sinusal Vías internodulares Nodo Aurículoventricular Haz AV Haces derecho e izquierdo de fibras de Purkinje.
Nódulo Sinoauricular
Esta es una banda aplanada de 3 x 15 x 1 mm, localizado en la pared posterolateral de la Aurícula derecha. Genera el impulso rítmico normal y posee fibras pequeñas musculares que casi no poseen filamentos musculares contráctiles. Están conectados directamente con las fibras musculares de las aurículas, por lo que el potencial de acción se propaga inmediatamente a la pared muscular auricular. La capacidad de autoexcitación se debe a la permeabilidad natural de la membrana a los iones de Na+ y Ca++ que hacen que el potencial en reposo sea menos negativo (-55 a -60mV). Debido a la poca negatividad en el interior de la membrana, los canales rápidos de Na+ se han inactivado debido a que las compuertas de inactivación se cierran y permanecen de esta manera.
La permeabilidad natural de las fibras nodulares hacia al Na+ permite el aumento progresivo del potencial de membrana hasta alcanzar el umbral (-40mV), los canales lentos de Na+ – Ca+ se activan, produciendo el potencial de acción. El término del potencial se debe a: •Cierre de los canales de Ca++ y Na+ después de un tiempo determinado (100- 150 ms) •Apertura de los canales de K+ La permeabilidad de la membrana al Ca++ y Na+ hace que la hiperpolarización se dé por menos tiempo, y haciendo que el potencial de membrana se desplace hacia arriba, alcanzando nuevamente el umbral de excitación.
Nodo Auriculoventricular
Existe un retraso en la conducción del impulso de las aurículas a los ventrículos, que permiten que las aurículas vacían sus contenidos en ventrículos antes de que comiencen su contracción. Un retraso total de .16 segundos hasta que llegue el impuso desde el nodo SA hasta las ramas del haz auriculoventricular. Esta conducción lenta se debe principalmente por la disminución del número de uniones en hendidura entre células sucesivas de las vías de conducción, de modo que hay una gran resistencia a la conducción de los iones excitadores desde una fibra de conducción hasta la siguiente.
Sistema de Purkinje
Este sistema posee fibras grandes (más grandes que las fibras musculares ventriculares. Transmiten el potencial rápidamente (1.5 a 4 m/s) → rapidez de propagación del impulso cardíaco. Se debe al alto nivel de permeabilidad de uniones en hendidura de los discos intercalares Tienen pocos elementos contráctiles, y esto hace que se contraigan poco o nada durante la transmisión del impulso. El haz AV es el único lugar donde no existen separación entre aurículas y ventrículos por una barrera fibrosa continua y esta barrera fibrosa actúa como un aislante. Como consecuencia la conducción del impulso por el haz AV es unidireccional (evita que el impulso vuelva a entrar a la aurícula). Transmisión del impulso cardíaco en el músculo ventricular El impulso viaja desde el endocardio hasta la superficie del corazón en un trayecto en espiral Tiempo total de trasmisión del impulso desde las ramas iniciales del haz AV hasta las últimas fibras musculares ventriculares es de 0.06 s.
Control de excitación y conducción en corazón
Todos los nodos y fibras del sistema cardionector poseen una tasa de descarga continua e independientemente, siendo la del nodo SA la mayor 70 a 80 veces por minuto. Nodo SA descarga con mucha mayor rapidez y emite un nuevo impulso antes que el nodo AV y las fibras de Purkinje puedan alcanzar su umbral de autoexcitación, por lo que se lo denomina el marcapasos normal del corazón. Frecuencia de descarga del nodo AV y fibras de Purkinje → nodo AV 40 a 60 veces por minuto; fibras de Purkinje 15 a 40 veces por minuto. Marcapasos ectópico → marcapasos en un lugar diferente del nodo SA. Bloqueo del nodo SA → Cuando se bloquea el nodo SA, el nodo AV asume frecuentemente el marcapasos Bloqueo del nodo AV → En aurículas asume nodo SA; en ventrículos asume las fibras de Purkinje (contracciones auriculares y ventriculares no son coordinadas). Sistema de Purkinje → Poco tiempo en la transmisión permite contracción sincronizada de los ventrículos, lo que aumenta la efectividad del latido.
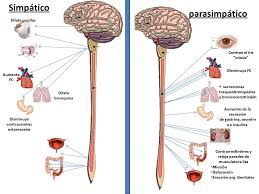
Sistema nervioso autónomo
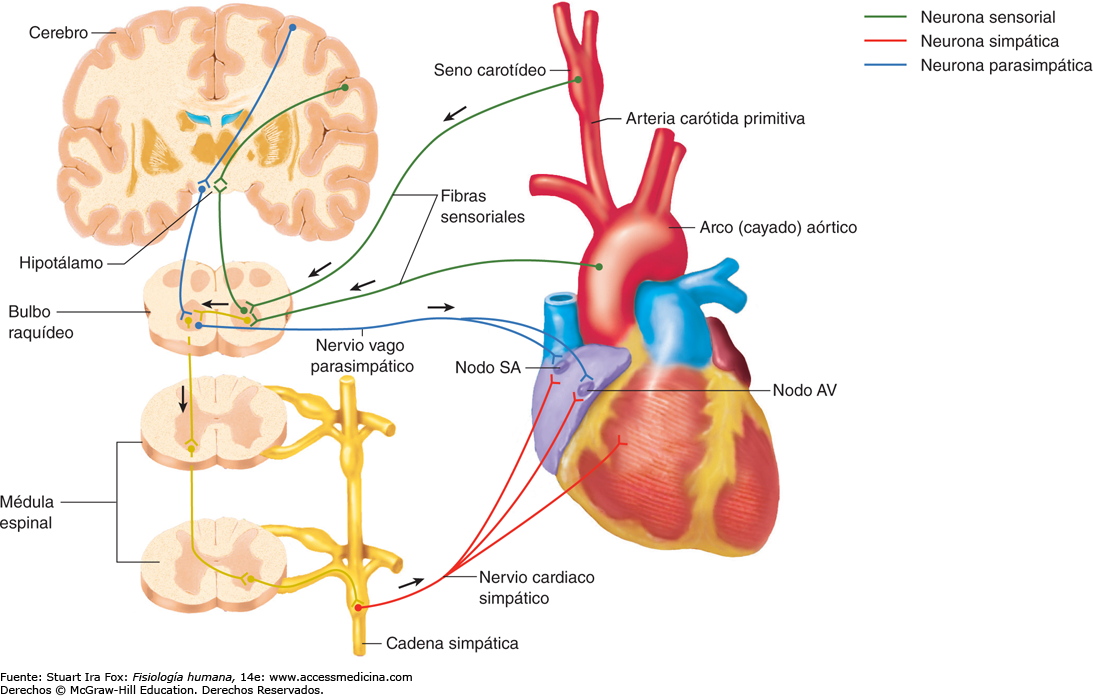
Parasimpático → Fibras se distribuyen por nodos SA y AV, y en menor medida en las fibras musculares auriculares y ventriculares. Su efecto disminuye la frecuencia del ritmo del nodo SA y también disminuye la excitabilidad de las fibras de unión AV. Debido a que la ACh aumenta la permeabilidad de K+ en la membrana causando la hiperpolarización (-65 a -75mV).
Fibras se distribuyen en todo el corazón, especialmente ventrículos. Su efecto aumenta la tasa de descarga del nodo SA, la tasa de conducción por el aumento de la excitabilidad y la fuerza de contracción. Esto se debe a que las catecolaminas aumentan la permeabilidad de la membrana al Na+ y Ca++.
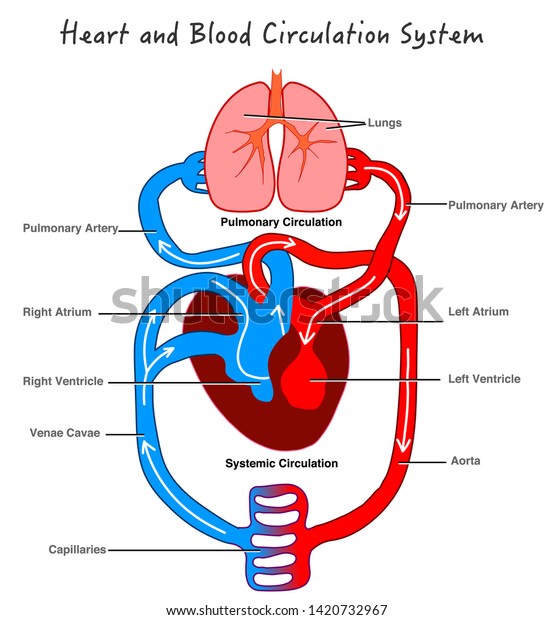
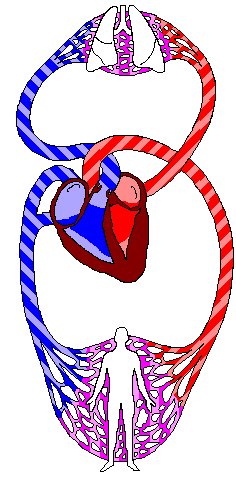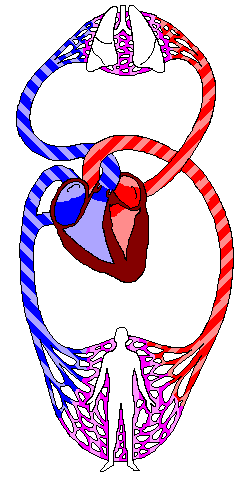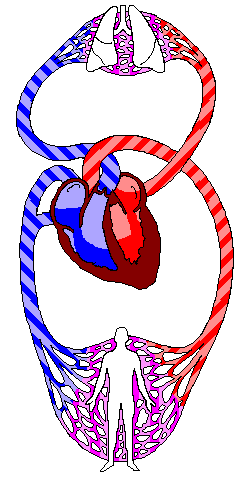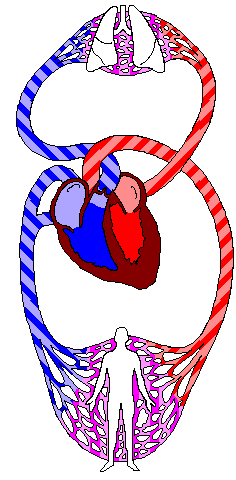
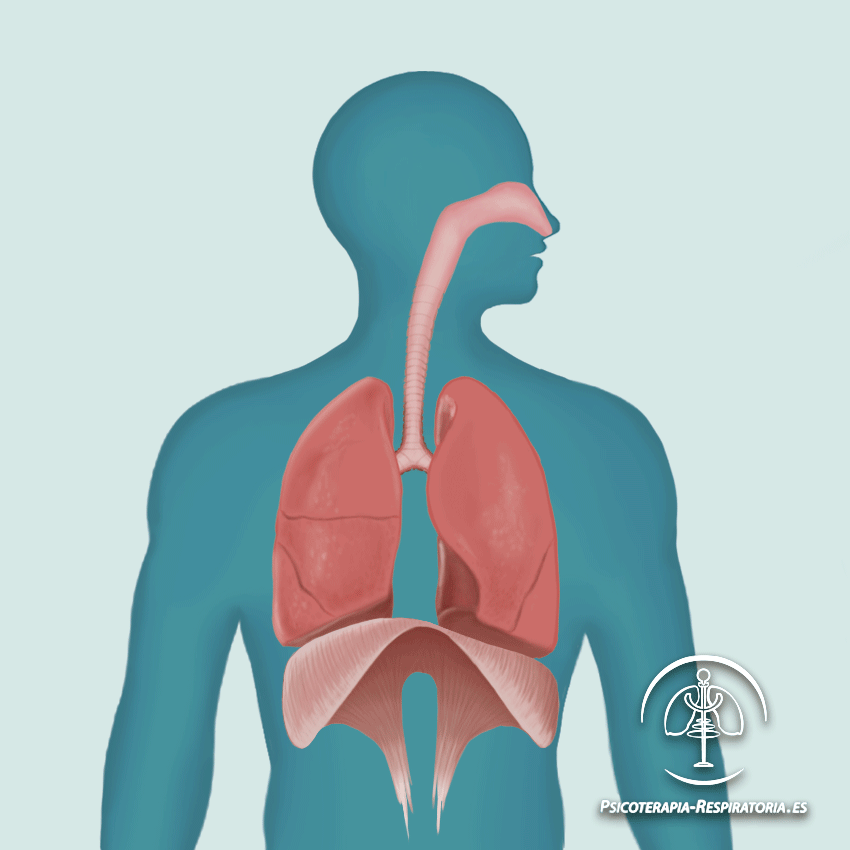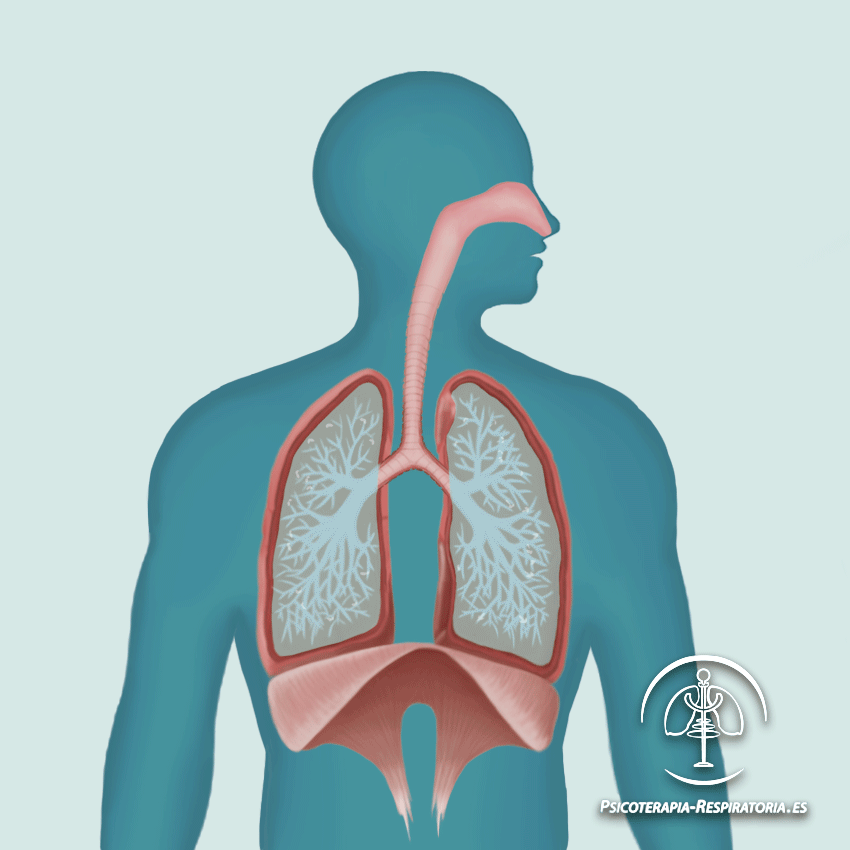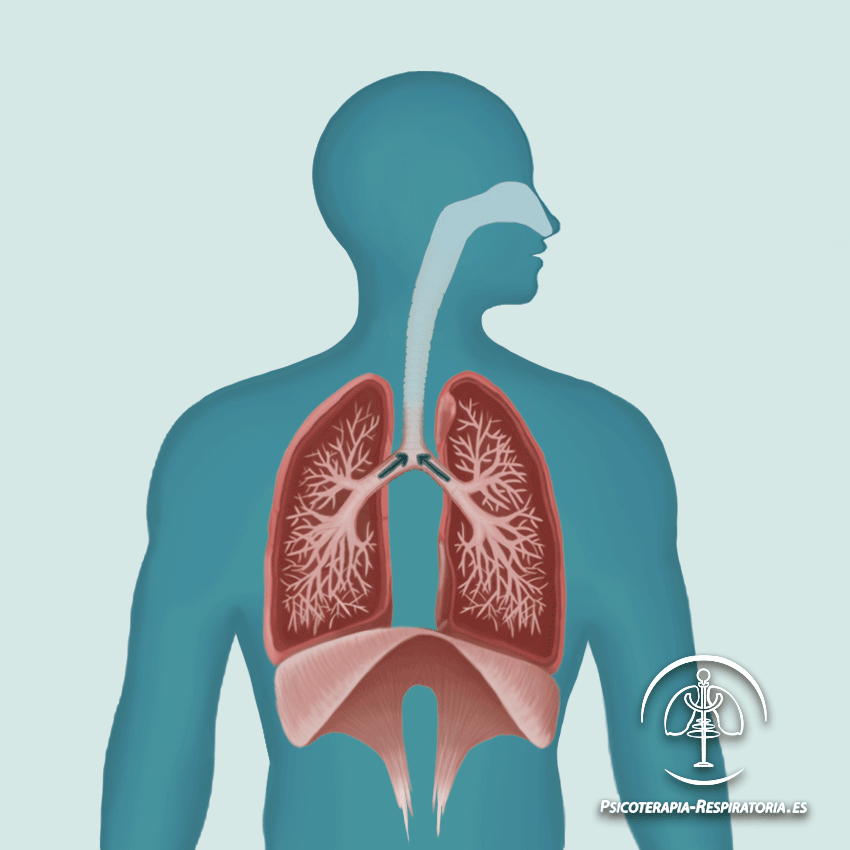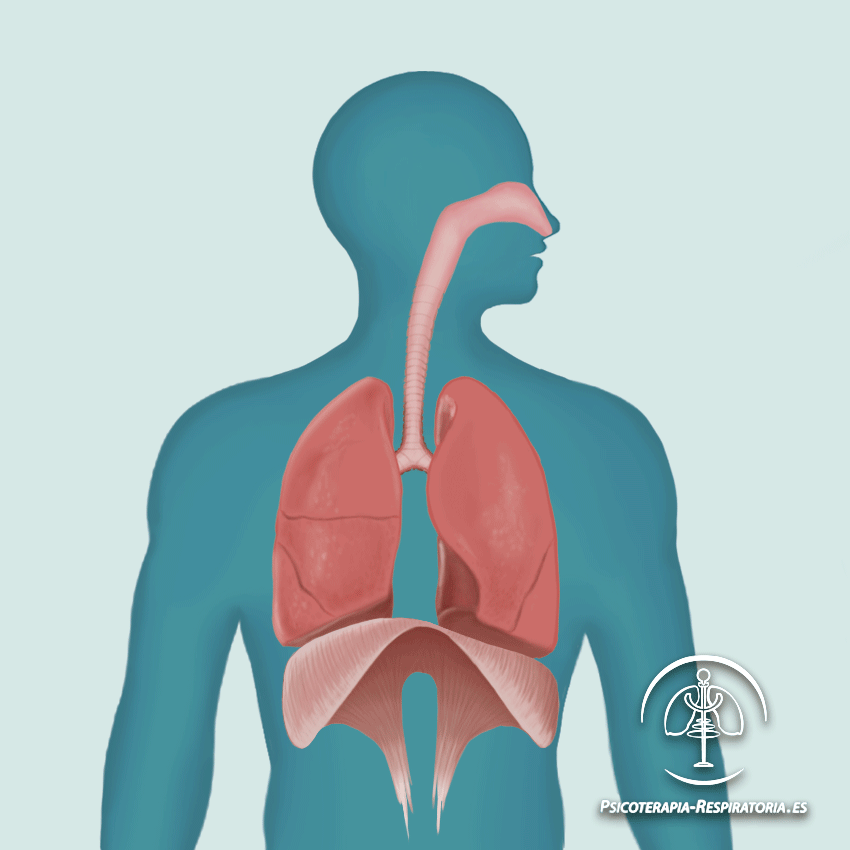
El corazón está formado por: por un Corazón derecho que bombea sangre a los pulmones y un Corazón izquierdo que bombea sangre a los tejidos periféricos, por medio de la circulación sistémica. Así mismo el corazón, está conformado por aurículas o atrios y por ventrículos. Una aurícula (que esta lleva la sangre hacia el ventrículo correspondiente) y un ventrículo (que da el impulso para que la sangre salga eyectada vaya a la circulación pulmonar o a todos los tejidos de nuestro cuerpo “circulación sistémica”).
Fisiología del músculo cardíaco
Está formado por tres tipos de músculos: • Músculo auricular • Músculo ventricular • Fibras musculares especializadas de excitación y de conducción. Entre las fibras musculares habrá discos intercalados, los cuales servirán para que los iones se muevan con facilidad en el líquido extracelular a lo largo de la fibra y así facilitar los potenciales de acción.
El corazón está formado por dos sincitios: el sincitio auricular que forma parte de las dos aurículas y el sincitio ventricular que forma parte de los ventrículos. Estos sincitios están separados por un tejido conectivo fibrosos dispuesto en forma de anillo, su función de esta estructura anatomica, sirve como un aislante electrico que hace que los potenciales de acción no se propaguen por el músculo si no por el HAZ AV. Permitiendo que los atrios se contraigan primero que los ventriculos, por un intervalo de tiempo de 0.1 segundos.
Potenciales de acción del musculo cardiaco
El potencial de acción del musculo cardiaco es producido por la apertura de dos tipos de canales: ✓Canales rápidos de sodio ✓canales lentos de calcio (canales de Ca+ – Na+) son estos los que orginan la meseta. ✓ Inmediatamente después del inicio del potencial de acción, la permeabilidad de la membrana del músculo cardiaco a iones K+ disminuye. Cuando se cierran los canales lentos de Ca+ es cuando empieza a salir el potasio y se vuelve a repolarizar la membrana. ✓ La velocidad de conducción en el sistema especializado de conducción del corazón, en las fibras de Purkinje que hay que entender y aclarar que no nos nervios son fibras musculares especializadas, es de hasta 4 m/s en la mayoría de las partes del sistema.
Periodo refractario y absoluto del musculo cardiaco
El periodo refractario normal del ventrículo es de 0.25 – 0.30 segundos. Este se extiende hasta la segunda mitad de la fase 3. Se puede excitar con una señal excitadora muy intensa El periodo refractario del musculo auricular es de 0.15 segundos aproximadamente. La duración de la contracción depende del potencial de acción, el musculo auricular es de 0,2 segundos y el musculo ventricular dura 0,3 segundos.
Ciclo cardiaco
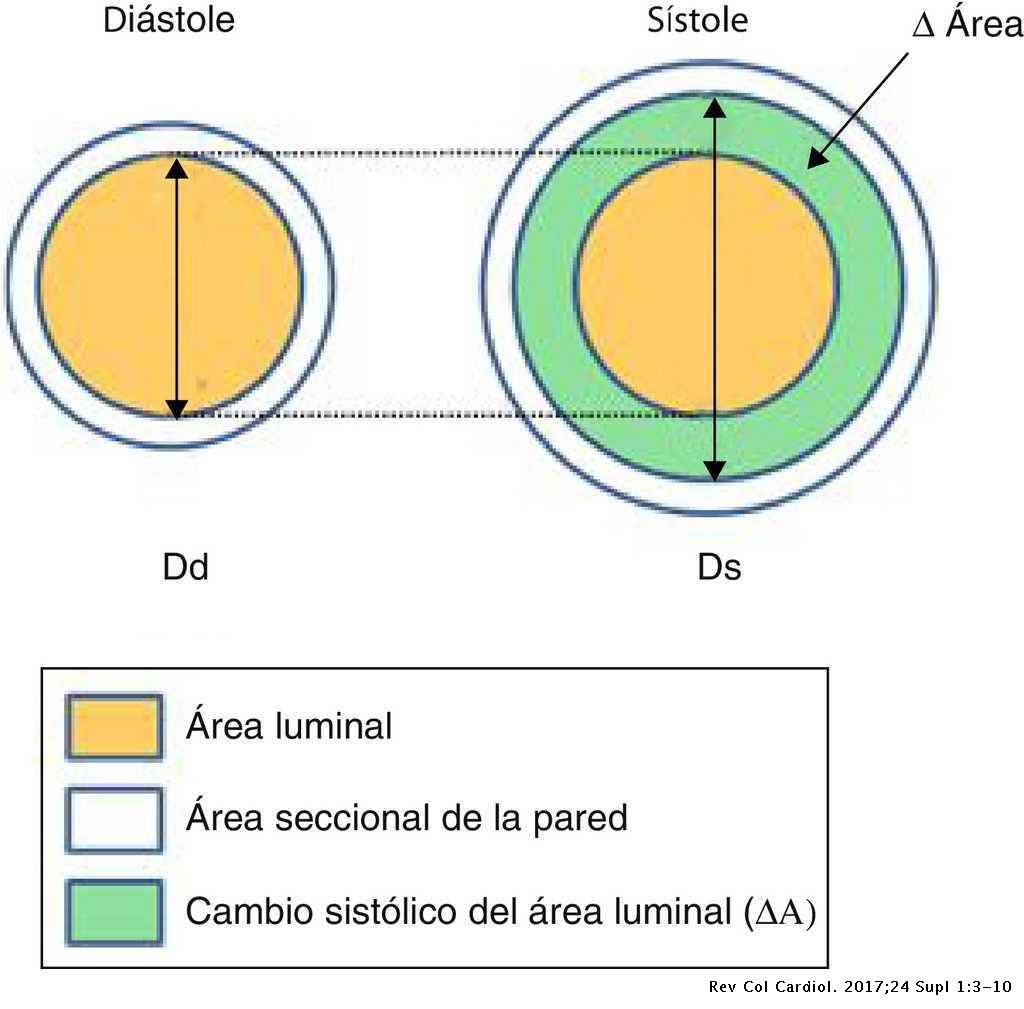
Se denomina ciclo cardíaco al conjunto de una sístole (el corazón se contrae expulsando la sangre que contienen los ventrículos hacia las arterias) y una diástole (al relajarse se vuelve a llenar de sangre). Periodo de contracción isovolumetrica: Desde que se cierran las válvulas AV hasta que se abren las Semilunares. El volumen de sangre intraventricular no varía, ya que al no estar abiertas las válvulas sigmoideas la sangre no puede ser expulsada en ninguna dirección.
Periodo de eyección ventricular: Comienza cuando la presión intraventricular supera la presión de la aorta y la arteria pulmonar, y se abren por ello las válvulas aórtica y pulmonar. Al principio es muy rápida y luego algo más lenta. En condiciones normales, la válvula aórtica se abre después y se cierra antes que la pulmonar.
Diástole ventricular: es el periodo de relajación durante el cual tiene lugar el llenado ventricular (período de repleción ventricular). La relajación de los ventrículos hace que disminuya la presión en su interior, por lo que al inicio de esta fase la presión en la aorta y en la arteria pulmonar supera la intraventricular, y esto hace que se cierren las válvulas aórtica y pulmonar. El cierre de las válvulas no es instantáneo, produciéndose cierta cantidad de reflujo (flujo negativo en la gráfica, que indica la inversión de su sentido). Consta de cuatro fases: Período de relajación isovolumétrica: Desde que se cierran las válvulas sigmoideas hasta que se abren las AV. En esta fase el volumen de sangre de los ventrículos no varía. Fase de llenado rápido: comienza cuando la presión intraventricular se hace inferior a la auricular, y se abre la válvula AV correspondiente. Se inicia el llenado ventricular. Fase de llenado lento (diástasis): el ventrículo se sigue llenando, pero de forma más lenta.
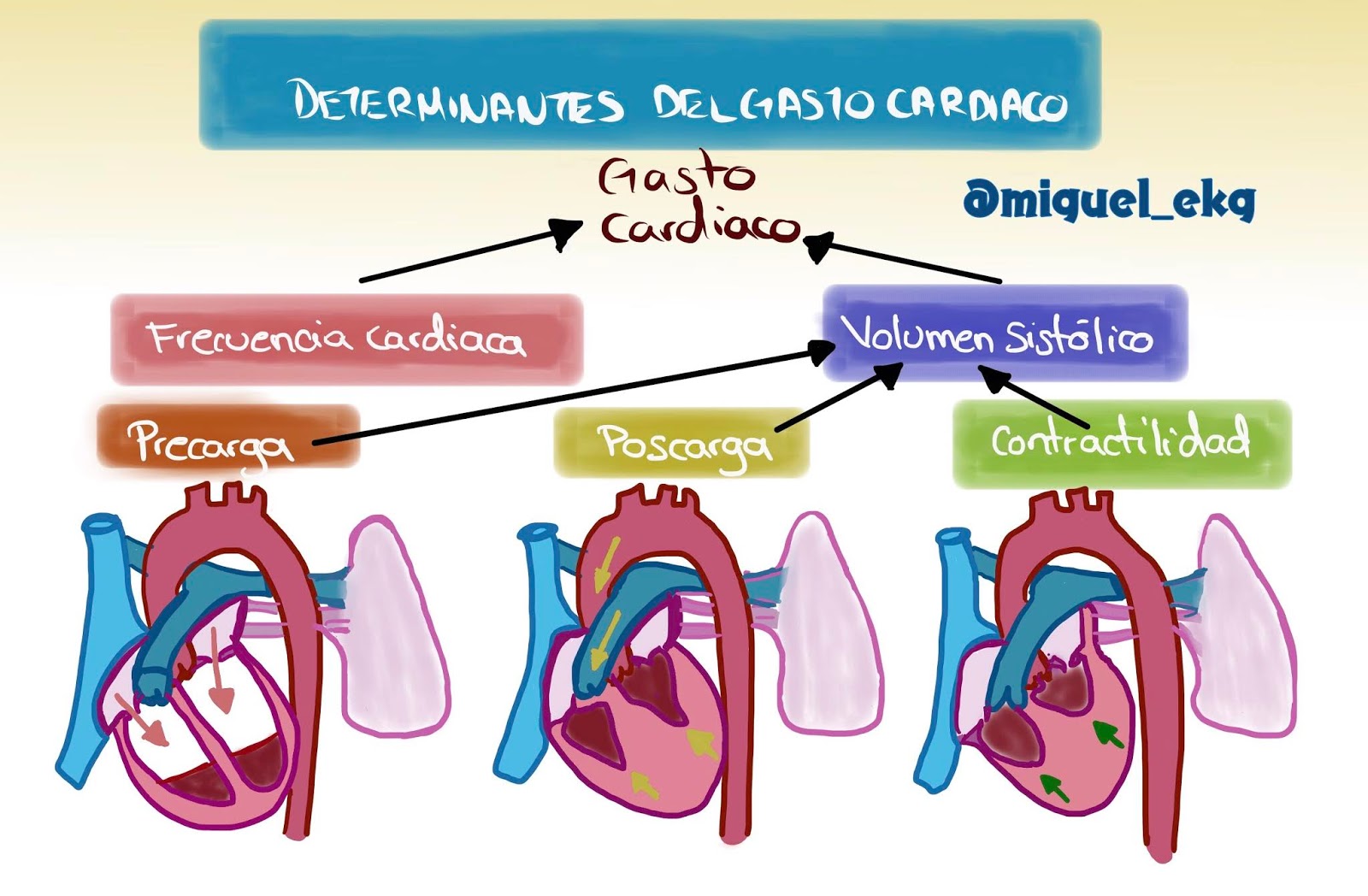
Regulación del bombeo cardiaco
En reposo, el corazón bombea de 4 a 6 litros de sangre por minuto, en una persona por decir estable o en reposo. Pero bombea mucho más cuando la persona está realizando ejercicio. Los mecanismos que van a controlar el volumen de bombeo del corazón son de acuerdo a dos mecanismos o formas: Regulación cardíaca intrínseca del bombeo o igual llamado Mecanismo de Frank-Starling. Por el sistema nervioso autónomo, en este caso el Simpático y el Parasimpático.
Vamos a hablar primero del primer mecanismo, vaya, por la cual la capacidad intrínseca del corazón va a adaptarse a volúmenes crecientes de flujo sanguíneo de entrada se denomina mecanismo de Frank-Starling del corazón.
Consiste en ver cuánto más se distiende el musculo cardiaco durante el llenado, mayor es la fuerza de contracción y mayor es la cantidad de sangre que bombea hacia la aorta (el corazón bombea toda la sangre que le llega procedente de las venas). En resumen y de manera mas sencilla si el corazón tiene una entrada de sangre pobre en oxígeno de 5,000 ml el corazón va a eyectar esos mismos 5,000 ml, el corazón va a sacar lo que le entre.
La eficacia de la función de bomba del corazón también está controlada por los nervios simpáticos (puede aumentar el gasto cardiaco a más del 100%) y parasimpáticos (vagos) (disminuye el gasto cardiaco casi a cero), que inervan de forma abundante el corazón.
Ejecutan acciones como la redistribución del flujo sanguíneo hacia las distintas zonas del organismo, el aumento o descenso de la actividad de bomba cardíaca y el control muy rápido de la presión arterial sistémica. El sistema nervioso controla la circulación casi totalmente a través del sistema nervioso autónomo.
Sistema nervioso autónomo
Cuenta con dos componentes:
Sistema nervioso simpático.
Sistema nervioso parasimpático.
La estimulación simpática causa vasoconstricción y aumenta la frecuencia y aumenta la frecuencia y la actividad de bombeo por el corazón
Los nervios salen de la médula espinal a través de los nervios de la columna torácica y algunas lumbares, se dirigen a las cadenas simpáticas para luego seguir dos rutas a la circulación: 1) a través de los nervios simpáticos que inervan la vasculatura de las vísceras y el corazón, y 2) en las porciones periféricas de los nervios raquídeos que se distribuyen a la vasculatura de las zonas periféricas. La mayoría de los vasos sanguíneos están inervados por fibras nerviosas simpáticas, a excepción de los capilares. La estimulación simpática aumenta la resistencia vascular y disminuye la velocidad del flujo sanguíneo.
La estimulación parasimpática disminuye la frecuencia y la actividad de bombeo por el corazón
Su principal función en el control de la circulación consiste en provocar un importante descenso de la frecuencia cardíaca y un pequeño descenso de la contractilidad del músculo cardíaco.
Control del sistema vasoconstrictor simpático por el sistema nervioso central
Centro vasomotor del cerebro y control del sistema vasoconstrictor. Situado bilateralmente en la sustancia reticular del bulbo y en el tercio inferior de la protuberancia, conforma una zona denominada centro vasomotor. Este centro transmite los impulsos parasimpáticos a través de los nervios vagos hacia el corazón y transmite los impulsos simpáticos a través de la médula espinal y los nervios simpáticos periféricos prácticamente hacia todas las arterias, arteriolas y venas del organismo.
El tronco simpático vasoconstrictor continuo provoca la construcción de la mayoría de los vasos sanguíneos. Este tono vasoconstrictor simpático mantiene un estado parcial de la contracción de los vasos sanguíneos. Cuando esta bloqueado por la anestesia raquídea los vasos se dilatan y el presión desciende hasta lo 50 mmHg.
Control del centro vasomotor por los centros nerviosos superiores
Áreas situadas en el mesencéfalo, sustancia reticular de la protuberancia y el diencéfalo, inhiben o excitan el centro vasomotor. El hipotálamo ejerce potentes reacciones en el centro vasomotor, tanto inhibidoras como excitadras. Partes de la corteza cerebral inhiben o excitan el centro vasomotor.
La noradrenalina es el neurotransmisor del sistema vasoconstrictor simpático.
Las sustancias de los nervios vasoconstrictores actúan directamente en los receptores a-adrenergéticos del músculo liso vascular, provocando vasoconstricción.
La médula suprarrenal libera noradrenalina y adrenalina durante la estimulación simpática
Ambas se transportan enbel torrente sanguíneo hacia todas las partes del organismo actuando sobre los vasos , provocando normalmente vasoconstricción al estimular receptores a-adrenérgicos, aunque la adrenalina posee efectos B-adrenérgicos, que provocan vasodilatación en algunos tejidos, como el músculo esquelético.
Función del sistema nervioso en el control rápido de la presión arterial
Una de las funciones más importante es su capacidad para provocar incrementos rápidos de presión arterial, provocando vasoconstricción y estimulación del corazón. Se producen tres cambios importante simultáneamente, que aumentan la presión al estimular el sistema nervioso autónomo:
Las venas y los demás vasos grandes de la circulación se contraen, lo que desplaza la sangre de los vasos periféricos al corazón y aumenta la fuerza de bombeo, lo que aumenta la fuerza de presión arterial.
El sistema nervioso autónomo estimula directamente el corazón, lo que potencia la bomba cardíaca.
La mayoría de lar arteriolas de la circulación sistémica se contraen, la que aumenta la resistencias vascular perfiérica total – presión arterial.
El sistema nervioso autónomo contribuye al aumento de la presión arterial durante el ejercicio intenso. Se sabe que aumenta de 30-40%. Al mismo tiempo que se activan las zonas motoras cerebrales para iniciar el ejercicio, se activa también la mayor parte del sistema activador reticular del tronco encefálico , incluye una mayor estimulación de zonas vasoconstrictoras y cardioaceleradoras del centro vasommotor. Es instanténeo el aumento de la presión para mantener la sincronización con el ejercicio intenso.
El sistema nervioso autónomo aumenta la presión arterial durante la reacción de alarma. Cuando se experimenta miedo intenso, la presión arterial aumenta hasta 200 mmHg en seg. Esta “reacción de alarma” proporciona aumento de la presión arterial que puede aportar sangre a cualquier músculo que necesite una respuesta instantánea para huir del peligro.
Mecanismos de reflejo para mantener la presión arterial normal
El sistema nervioso autónomo actúa para mantener la presión arterial en valores prácticamente normales a través de mecanismos reflejos de retroalimentación negativa.
Sistema de control de la presión arterial mediante barorreceptores
Básicamente, este reflejo se inicia en los receptores de estiramiento, conocidos como barorreceptores o presorreceptores, situados en puntos específicos de las paredes de varias arterias sistémicas de gran tamaño. Los barorreceptores son muy abundantes en: 1) la pared de ambas arterias carótidas internas, a corta distancia por encima de la bifurcación carotídea (una zona que se conoce como seno carotídeo), y 2) en la pared del cayado aórtico. Las señales de los «barorreceptores carotídeos» se transmiten a través de los pequeños nervios de Hering, hacia los nervios glosofaríngeos de la parte alta del cuello y después hacia el tracto solitario de la zona del bulbo en el tronco del encéfalo. Las señales que proceden de los «barorreceptores aórticos» del cayado aórtico se transmiten a través de los nervios vagos también hacia el tracto solitario del bulbo.
Los barorreceptores sinusales carotídeos no se estimulan en absoluto con presiones entre 50-60 mmHg, pero en valores superiores responden con una frecuencia progresivamente mayor y alcanzan el máximo en torno a los 180 mmHg. Los barorreceptores responden mucho más a una presión que cambia con gran rapidez que a una presión estacionaria.
Reflejo circulatorio iniciado por los barorreceptores
Después de que las señales de los barorreceptores entren en el tracto solitario del bulbo, las señales secundarias inhiben el centro vasoconstrictor del bulbo y excitan el centro parasimpático vagal. Los efectos netos son dos; 1) la vasodilatación de las venas y arteriolas en todo el sistema circulatorio periférico y 2) el descenso de la frecuencia cardíaca y de la fuerza de contracción cardíaca.
Función de los barorreceptores durante los cambios de postura del cuerpo
La capacidad de los barorreceptores de mantener una presión arterial relativamente constante en la parte superior del cuerpo es importante cuando una persona se levanta después de haber estado tumbada.
Función «amortiguadora» de la presión del sistema de control de barorreceptores
Como el sistema de barorreceptores se opone tanto al aumento como al descenso de la presión arterial, se denomina sistema amortiguador de la presión y los nervios de los barorreceptores se conocen como nervios amortiguadores. Uno de los objetivos principales del sistema arterial de barorreceptores consiste en reducir minuto a minuto la variación de la presión arterial hasta un tercio de la que aparecería si no estuviera presente este sistema.
¿Son importantes los barorreceptores en la regulación a largo plazo de la presión arterial?
Aunque los barorreceptores arteriales proporcionan un control potente de la presión arterial minuto a minuto, su importancia en la regulación a largo plazo de la presión sanguínea es controvertida como consecuencia, tal vez, de que algunos fisiólogos consideran que los barorreceptores tienen una importancia relativamente escasa en la regulación crónica de la presión arterial, porque tienden a reajustarse en 1-2 días a la presión a la cual se exponen.
Control de la presión arterial por los quimiorreceptores carotídeos y aórticos: efecto de la falta de oxígeno sobre la presión arterial
Estrechamente asociado al control de los barorreceptores del sistema de presión actúa un reflejo de quimiorreceptores que funciona de una forma muy similar al reflejo de barorreceptores, excepto porque son los quimiorreceptores, y no los receptores de estiramiento, los que inician la respuesta. Están formados por células quimiosensibles a la ausencia de oxígeno, al exceso de dióxido de carbono y al exceso de iones hidrógeno. Se localizan en dos cuerpos carotídeos, cada uno de los cuales se sitúa en la bifurcación de cada arteria carótida común, y habitualmente entre uno y tres cuerpos aórticos adyacentes a la aorta. Excitan fibras nerviosas que llegan por los nervios de Hering y los nervios vagos hacia el centro vasomotor del tronco del encéfalo. Siempre que la presión arterial cae por debajo de un nivel crítico los quimiorreceptores se estimulan porque el descenso del flujo sanguíneo provoca la disminución del oxígeno y también la acumulación excesiva de dióxido de carbono e iones hidrógeno que no se eliminan por una sangre que fluye lentamente. Las señales transmitidas desde los quimiorreceptores excitan el centro vasomotor, lo que eleva la presión arterial hasta la normalidad
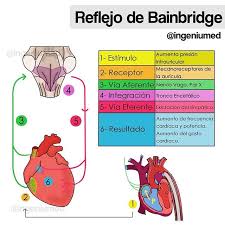
Control del reflejo auricular de la frecuencia cardíaca (reflejo Bainbridge)
El aumento de la presión auricular también aumenta la frecuencia cardíaca, a veces hasta en un 75%. Una pequeña parte de este incremento se debe al efecto directo del aumento del volumen auricular para estirar el nódulo sinusal. Otro 40-60% del aumento de la frecuencia de un reflejo nervioso denominado reflejo de Bainbridge. Los receptores de estiramiento de las aurículas que provocan el reflejo de Bainbridge transmiten sus señales aferentes a través los nervios vagos hacia el bulbo raquídeo. Sirve para prevenir el estancamiento de la sangre en las venas, las aurículas y la circulación pulmonar.
Respuesta isquémica del sistema nervioso central: control de la presión arterial por el centro vasomotor del cerebro en respuesta a un descenso del flujo sanguíneo cerebral
Cuando el flujo sanguíneo que se dirige hacia el centro vasomotor en la parte inferior del tronco del encéfalo disminuye lo suficiente para provocar un defecto nutricional, es decir, para provocar la isquemia cerebrales neuronas vasoconstrictoras y cardioaceleradoras del centro vasomotor responden directam ente a la isquemia y se excitan con fuerza. Esta elevación en respuesta a una isquemia cerebral se conoce como la respuesta isquémica del sistema nervioso central (SNC).
El grado de vasoconstricción simpática provocado por la isquemia cerebral intensa a menudo es tan grande que algunos de los vasos periféricos se ocluyen total o casi totalmente. Por tanto, la respuesta isquémica del SNC es uno de los activadores más potentes de todos los activadores del sistema vasoconstrictor simpático.
Reacción de Cushing al aumento de la presión en torno al encéfalo
La denominada reacción de Cushing es un tipo especial de respuesta isquémica del SNC que se produce como consecuencia del aumento de presión del líquido cefalorraquídeo que rodea al cerebro en la bóveda craneal. Protege a los centros vitales del cerebro de la pérdida de nutrientes en caso de que la presión del líquido cefalorraquídeo sea suficientemente alta para comprimir las arterias cerebrales.
Control local del flujo sanguíneo en respuesta a las necesidades tisulares
Uno de los principios de la circulación consta en que los tejidos controlan su propio flujo sanguíneo en proporciona a sus necesidades, las cuales son:
1.- Aporte de oxigeno a los tejidos. 2.- Aporte de otros nutrientes, como glucosa, aminoácidos y ácidos grasos. 3.-Eliminaciones de dióxido de carbono de los tejidos. 4.- Eliminaciones de iones hidrógeno de los tejidos. 5.- Mantenimientos de las concentraciones adecuadas de otros iones en los tejidos. 6.- Transporte de varias hormonas y otras sustancias a los distintos tejidos.
Variaciones del flujo sanguíneo en distintos tejidos y órganos
Un flujo sanguíneo total de 1.350 ml/min en el hígado, que es de 95 mi/ min/lOOg de tejido hepático; el flujo sanguíneo tan importante que atraviesa los riñones, 1.100ml/min; en los tejidos el flujo sanguíneo siempre está controlado a un nivel que solo es ligeramente mayor de lo necesario para mantener la oxigenación tisular plena; es sorprendente el escaso flujo sanguíneo que llega a todos los músculos inactivos del organismo, sólo un total de 750 ml/min; durante el ejercicio intenso la actividad metabólica muscular aumenta más de 60 veces y el flujo sanguíneo hasta 20 veces, aumentando hasta 16.000 m l/min en el lecho vascular muscular total del cuerpo.
Mecanismos de control del flujo sanguíneo
El control del flujo sanguíneo local se puede dividir en:
1) Control a corto plazo. 2) Control a largo plazo.
El control a corto plazo se consigue con cambios rápidos de la vasodilatación o vasoconstricción local de las arteriolas, metaarteriolas y esfínteres precapilares, que se producen en segundos o minutos para proporcionar con gran rapidez el mantenimiento del flujo sanguíneo tisular local apropiado
El control a largo plazo significa cambios controlados lentos del flujo en un período de días, semanas o incluso meses.
Regulación a corto plazo del flujo sanguíneo local cuando cambia la disponibilidad de oxígeno
El envenenamiento por cianuro en un territorio altera el uso de oxígeno provocando un aumento local del flujo sanguíneo hasta de siete veces, lo que demuestra el efecto extremo que tiene la deficiencia de oxígeno para aumentar el flujo sanguíneo. Hay dos teorías básicas para la regulación del flujo sanguíneo local cuando cambia el metabolismo tisular o disponibilidad de oxígeno: 1) la teoría vasodilatadora y 2) la teoría de la falta de oxígeno.
Teoría vasodilatadora de la regulación a corto plazo del flujo sanguíneo local: posible papel especial de la adenosina
Cuanto mayor sea el metabolismo o menor sea la disponibilidad de oxígeno o de algunos otros nutrientes en un tejido, mayor será la velocidad de formación de sustancias vasodilatadoras, las cuales se difunden a través de los tejidos hacia los esfínteres precapilares, las metaarteriolas y las arteriolas para provocar la dilatación, ejemplos de estas son: adenosina, dióxido de carbono, compuestos con fosfato de adenosina, histamina, iones potasio e iones hidrógeno.
Teoría de la falta de oxígeno
Con mayor exactitud, la teoría de la falta de nutrientes (porque están implicados otros nutrientes, además del oxigeno). El oxigeno (y también otros nutrientes) es necesario como uno de los nutrientes metabólicos para provocar la concentración muscular. Por lo tanto es razonable creer que los vasos sanguíneos simplemente se relajarían en ausencia de una cantidad adecuada de oxigeno, dilatándose de forma natural. Además el aumento del metabolismo podría en teoría disminuir la disponibilidad de oxigeno hacia las fibras musculares lisas de los vasos sanguíneos locales, lo cual también provocaría la vasodilatación local.
Los esfínteres precapilares y las metaarteriolas se abren y cierran cíclicamente varias veces por minuto, siendo proporcional la duración de las fases abiertas a las necesidades metabólicas de oxígeno en los tejidos. La apertura y el cierre cíclicos se denominan vasomotilidad.
Ejemplos especiales del control «metabólico» a corto plazo del flujo sanguíneo local
Hiperemia reactiva. Cuando la sangre que irriga un tejido se bloquea durante unos segundos durante lh o más, y después se desbloquea, el flujo sanguíneo que atraviesa el tejido aumenta inmediatamente hasta 4-7 veces con respecto a lo normal; este aumento del flujo continuará durante varios segundos, si el bloqueo ha durado sólo unos segundos, pero a veces continuará muchas horas, si el flujo sanguíneo ha estado interrumpido durante lh o más. Este fenómeno se conoce como hiperemia reactiva. Resalta la estrecha conexión existente entre la regulación del flujo sanguíneo local y el aporte de oxígeno y de otros nutrientes a los tejidos.
Hiperemia activa. Cuando cualquier tejido se vuelve muy activo, como un músculo que hace ejercicio, una glándula gastrointestinal durante el período hipersecretor o incluso en el cerebro durante la actividad mental rápida, la velocidad del flujo sanguíneo aumenta a través del tejido. El incremento del metabolismo local hace que las células devoren rápidamente los nutrientes del líquido tisular y también que liberen grandes cantidades de sustancias vasodilatadoras.
«Autorregulación» del flujo sanguíneo cuando la presión arterial cambia de la normalidad: mecanismos «metabólicos» y «miógenos»
En cualquier tejido del organismo el rápido incremento de la presión arterial provoca un aumento inmediato del flujo sanguíneo, pero en menos de l/min ese flujo volverá a la normalidad en la mayoría de los tejidos, incluso aunque la presión arterial se mantenga elevada, se denomina autorregulación.
Durante casi un siglo se han mantenido dos opiniones que explicarían el mecanismo de autorregulación a corto plazo, la teoría metabólica y la teoría miógena.
Teoría metabólica: Cuando la presión arterial es demasiado elevada, el exceso de líquido proporciona demasiado oxígeno y demasiados nutrientes de otro tipo hacia los tejidos y «lava» los vasodilatadores liberados por los tejidos. Estos nutrientes (en especial, el oxígeno), junto con el descenso en los niveles tisulares de vasodilatadores, provocan entonces la constricción de los vasos sanguíneos y el retorno del flujo casi a la normalidad, a pesar de que aumente la presión.
Teoría miógena: Se basa en la observación de que el estiramiento brusco de los vasos sanguíneos pequeños provoca la contracción del músculo liso de la pared vascular durante unos segundos. Se ha propuesto que cuando una presión arterial elevada estira el viso se provoca, a su vez, una constricción vascular reactiva reduce el flujo sanguíneo casi a la normalidad. Por el contrario, con presiones bajas el grado de estiramiento del vaso es menor, por lo que el músculo liso se relaja, reduce la resistencia vascular y ayuda a recuperar la normalidad del flujo.
Regulación a largo plazo del flujo sanguíneo
La mayoría de los mecanismos de regulación del flujo sanguíneo local que hemos comentado actúan en pocos segundos o minutos después del cambio de la situación tisular local. A pesar de ello, el flujo sanguíneo se ajusta solo en las tres cuartas partes de las necesidades adicionales de los tejidos, incluso después de la activación completa de esos mecanismos agudos. La regulación a largo plazo del flujo sanguíneo es especialmente importante cuando cambian las demandas metabólicas del tejido a largo plazo. Es decir, si un tejido esta crónicamente hiperactivo y, por tanto requiere u aumento crónico de las cantidades de oxigeno y otros nutrientes, por lo que en alguna semanas aumentan tanto el numero como el tamaño de las arteriolas y los vasos capilares para cubrir las necesidades del tejido, a menos que el aparato circulatorio se vuelva patológico o sea demasiado viejo para responder.
Las arterias y arteriolas muy pequeñas de cada tejido mediante la deficiencia de sustancias vasodilatadoras o de oxigeno en las células tisulares solo pueden llegara esos vasos, no a las arterias intermedias y mayores del territorio proximal. Aun así, cuando aumenta el flujo sanguíneo a través de una porción microvascular de la circulación, secundariamente se activa otro mecanismo que dilata también las arterias mayores sintetizan varias sustancias que liberadas afectan el grado de relajación o contracción de la pared arterial.
El flujo de sangre rápido a través de las arterias y arteriolas provoca fuerzas de cizallamiento sobre las células endoteliales.
Función del oxigeno en la regulación a largo plazo
El oxigeno es importante no solo para el control a corto plazo del flujo sanguíneo local, sino también para el control a largo plazo. Un ejemplo es el aumento de la vascularización de los tejidos en los animales que viven en altitudes elevadas, donde el oxigeno atmosférico es bajo. El exceso de oxigeno provoca la interrupción casi inmediata del crecimiento vascular nuevo en la retina de los ojos del niño prematuro incluso provoca la degeneración de algunos de los vaso pequeños que ya se han formado.
Hay una docena o más factores que aumentan el crecimiento de los vasos sanguíneos nuevos, siendo casi todos ellos péptidos pequeños, tres de los mejor identificados son el factor de crecimiento de los fibroblastos, el factor decrecimiento del endotelio vascular (VEGF) y la angiogenina, aislados cada uno de ellos en tejido que tienen un aporte sanguíneo inadecuado. Presumiblemente, es la deficiencia de oxigeno tisular o otros nutrientes la que provoca la formación de los factores de crecimiento vascular (también denominados “factores angiogenicos”)
Cuando se bloquean una arteria o una vena en cualquier tejido del organismo se desarrolla un canal vascular nuevo rodeando el bloqueo y permitiendo que se vuelva a suministrar sangre al tejido afectado, al menos parcialmente. La primera etapa de este proceso es la dilatación de los bucles vasculares pequeños que ya conectan ese vaso proximal al bloqueo con el vaso distal. Después el flujo es menor de la cuarta parte de lo necesario para cubrir todas las necesidades tisulares.
Control humoral
Se refiere al control por las sustancias segregadas o absorbidas en los líquidos del organismo, como hormonas y factores producidos localmente.
Sustancias vasoconstrictoras
Noradrenalina y adrenalina. La noradrenalina es una hormona vasoconstrictora especialmente potente; la adrenalina es menos potente y en algunos tejidos provoca incluso una vasodilatación leve. Estas hormonas circulan entonces por todo el cuerpo y provocan casi los mismos efectos en la circulación que la estimulación simpática directa, con lo que se consigue un sistema de control doble: 1) estimulación nerviosa directa y 2) efectos indirectos de la noradrenalina y/o de la adrenalina; en la sangre circulante.
Angiotensina II. La angiotensina II es otra sustancia vasoconstrictora potente. Tan sólo una millonésima de gramo puede aumentar la presión arterial de un ser humano en 50 mmHg o más. El efecto de angiotensina II contrae potentemente las pequeñas arteriolas. Normalmente actúa sobre muchas de las arteriolas del organismo al mismo tiempo, para aumentar la resistencia periférica total y aumentar la presión arterial.
Vasopresina. La vasopresina, que también se conoce como hormona antidiurética. Se forma en las células nerviosas del hipotálamo, pero después se transporta distalmente a través de los axones nerviosos hacia la neurohipófisis, donde es finalmente segregada a la sangre. Tiene una función im portante aum entando la reabsorción de agua de los túbulos renales hacia la sangre y, por tanto, ayudando a controlar el volumen de líquido corporal.
Sustancias vasodilatadoras.
Bradicinina
Histamina
Control vascular por iones y otros factores químicos
Hay muchos iones y otros factores químicos que pueden dilatar o contraer los vasos sanguíneos locales.
La mayoría de ellos tiene una función escasa en la regulación global de la circulación, pero hay algunos efectos específicos, como son: 1. El aumento de la concentración del ion calcio provoca vasoconstricción. 2. El aumento de la concentración del ion potasio provoca vasodilatación. 3. El aumento de la concentración del ion magnesio provoca una vasodilatación potente. 4. El aumento de la concentración del ion hidrogeno (descenso de pH) provoca la dilatación de las arteriolas. 5. Los iones acetato y citrato, que provocan una vasodilatación pequeña. 6. El aumento dela concentración de dióxido de carbono provoca una vasodilatación moderada y la mayoría delos tejidos, pero una vasodilatación importante en el cerebro.
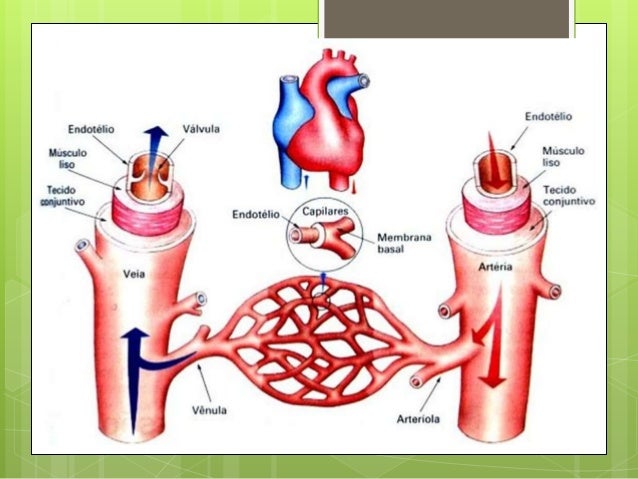
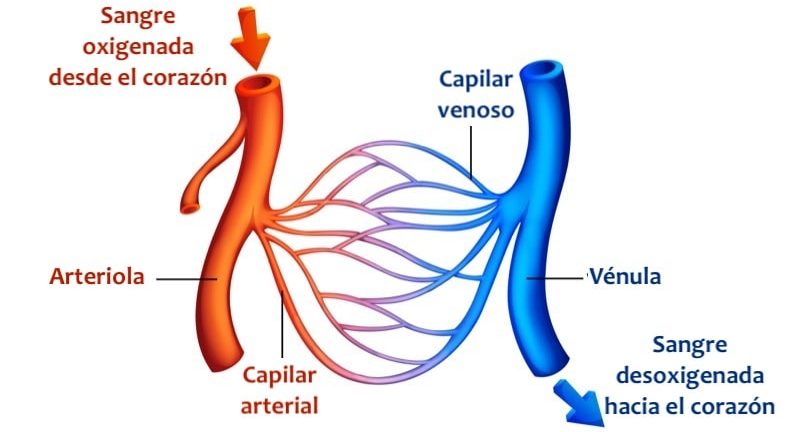
El principal objetivo de la microcirculación es el transporte de nutrientes hacia los tejidos y la eliminación de los restos celulares.
Las arteriolas pequeñas controlan el flujo sanguíneo hacia cada tejido y, a su vez, las condiciones locales de los tejidos controlan los diámetros de las arteriolas; es decir, que cada tejido controla, en la mayoría de los casos, su propio flujo sanguíneo dependiendo de sus necesidades individuales.
ESTRUCTURA DE LA MICROCIRCULACIÓN Y DEL SISTEMA CAPILAR
La microcirculación de cada órgano está organizada para atender sus necesidades específicas.
En general, cada arteria nutricia que entra en un órgano se ramifica seis u ocho veces antes de que las arterias sean suficientemente pequeñas para denominarse arteriolas, que, en general, tienen diámetros internos de solo 10-15 μm.
Entonces las arteriolas se ramifican entre dos y cinco veces, alcanzando diámetros de 5 a 9 μm en sus extremos cuando aportan la sangre a los capilares.
En el punto en el que cada capilar verdadero se origina de una metaarteriola hay una fibra muscular lisa que rodea el capilar. Esta estructura se conoce como esfínter precapilar. Este esfínter abre y cierra la entrada al capilar.
Las vénulas son mayores que las arteriolas y tienen una capa muscular mucho más débil. A pesar de ello, la presión de las vénulas es mucho menor que la de las arteriolas, por lo que las vénulas aún pueden contraerse considerablemente, a pesar de su capa muscular débil.
Esta distribución típica del lecho capilar no se encuentra en todas las partes del cuerpo, aunque algunas distribuciones similares pueden servir para el mismo objetivo.
ESTRUCTURA DE LA PARED CAPILAR
La estructura ultramicroscópica de las células endoteliales típicas de la pared capilar como se ven en la mayor parte de los órganos del cuerpo, en especial en los músculos y el tejido conjuntivo.
La pared está compuesta por una capa unicelular de células endoteliales y rodeada por una membrana basal muy fina en el exterior del capilar.
El grosor total de la pared capilar es de solo unas 0,5 μm, el diámetro interno del capilar es de 4-9 μm, apenas suficiente para el paso de los eritrocitos y otras células sanguíneas exprimidas.
POROS EN LA MEMBRANA CAPILAR
La figura muestra dos pequeños pasadizos que conectan el interior del capilar con el exterior. Uno de los pasos es un espacio intercelular, un canal curvo a modo de hendidura fina que descansa en la parte superior de la figura entre células endoteliales adyacentes.
Cada espacio está interrumpido periódicamente por pliegues cortos de inserciones de proteínas que mantienen unidas las células endoteliales, pero entre esos pliegues puede filtrarse libremente el líquido a través del espacio. El espacio suele tener un tamaño uniforme, con una anchura de 6-7 nm (60-70 angström), que es algo menor que el diámetro de una molécula de albúmina.
TIPOS ESPECIALES DE POROS EN LOS CAPILARES DE ALGUNOS ÓRGANOS
Los «poros» de los capilares de algunos órganos tienen unas características especiales para cumplir las necesidades peculiares de los órganos. Algunas de estas características son las siguientes:
1. En el cerebro, las uniones entre las células endoteliales capilares son principalmente uniones «estrechas» que permiten la entrada y salida de moléculas muy pequeñas como agua, oxígeno y dióxido de carbono en los tejidos cerebrales.
2. En el hígado sucede lo contrario. Los espacios entre las células endoteliales capilares son aperturas amplias, por lo que casi todas las sustancias disueltas en el plasma, incluidas las proteínas plasmáticas, pueden pasar de la sangre a los tejidos hepáticos. 3. Los poros de las membranas capilares gastrointestinales tienen un tamaño intermedio entre las de los músculos y las del hígado.
4. En los capilares glomerulares del riñón se abren numerosas membranas ovales, denominadasfenestraciones, que atraviesan en todo su trayecto las células endoteliales, por lo que pueden filtrarse cantidades enormes de moléculas pequeñas e iones (pero no las moléculas grandes de las proteínas plasmáticas) a través de los glomérulos sin tener que pasar a través de los espacios situados entre las células endoteliales.
FLUJO DE SANGRE EN LOS CAPILARES (VASOMOTILIDAD)
La sangre no fluye continuamente a través de los capilares, sino que lo hace de forma intermitente apareciendo y desapareciendo cada pocos segundos o minutos. La causa de esta intermitencia es el fenómeno conocido como vasomotilidad, lo que significa la contracción intermitente de las metaarteriolas y esfínteres precapilares (y, a veces, también de las arteriolas muy pequeñas).
REGULACIÓN DE LA VASOMOTILIDAD
El factor más importante que afecta al grado de apertura y cierre de las metaarteriolas y de los esfínteres precapilares, y que se ha descubierto hasta la fecha, es la concentración de oxígeno en los tejidos.
Cuando la velocidad de utilización del oxígeno por el tejido es mayor, de forma que la concentración de oxígeno tisular disminuye por debajo de lo normal, se activan los períodos intermitentes del flujo sanguíneo capilar más a menudo y la duración de cada período del flujo es más prolongada, con lo que se permite que la sangre capilar transporte mayores cantidades de oxígeno (y de otros nutrientes) hacia los tejidos.
FUNCIÓN MEDIA DEL SISTEMA CAPILAR
A pesar de que el flujo sanguíneo a través de cada capilar es intermitente, hay tantos capilares en los tejidos que su función global termina por ser superada, es decir, hay una velocidad media del flujo sanguíneo a través de cada lecho capilar tisular, una presión capilar media dentro de los capilares y una velocidad de transferencia media de las sustancias entre la sangre de los capilares y el líquido intersticial circundante.
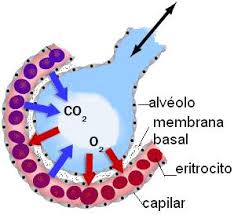
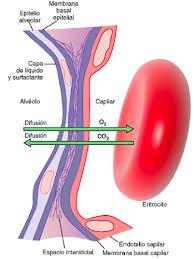
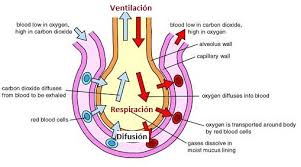
DIFUSIÓN A TRAVÉS DE LA MEMBRANA CAPILAR
Con mucho, el medio más importante por el cual se transfieren las sustancias entre el plasma y el líquido intersticial es la difusión. En la figura se ilustra este proceso, es decir, se ve cómo el flujo sanguíneo recorre la luz del capilar y la gran cantidad de moléculas de agua y partículas disueltas que entran y salen a través de la pared capilar, permitiendo la mezcla continua entre el líquido intersticial y el plasma.
La difusión es consecuencia del movimiento térmico de las moléculas de agua y de otras sustancias disueltas en el líquido, con las distintas moléculas e iones desplazándose primero en una dirección y luego en otra, rebotando aleatoriamente en cada una de ellas.
LAS SUSTANCIAS LIPOSOLUBLES DIFUNDEN DIRECTAMENTE A TRAVÉS DE LAS MEMBRANAS CELULARES DEL ENDOTELIO CAPILAR
Si una sustancia es liposoluble, difunde directamente a través de las membranas celulares del capilar sin tener que atravesar los poros. Estas sustancias son el oxígeno y el dióxido de carbono.
Como estas sustancias pueden atravesar todas las zonas de la membrana capilar, sus velocidades de transporte a través de la membrana capilar son muchas veces más rápidas que las de las sustancias insolubles en lípidos, como los iones sodio y la glucosa, que solo pueden pasar a través de los poros.
LAS SUSTANCIAS HIDROSOLUBLES Y NO LIPOSOLUBLES DIFUNDEN SOLO A TRAVÉS DE LOS POROS INTERCELULARES EN LA MEMBRANA CAPILAR
Muchas sustancias que necesitan los tejidos son solubles en agua pero no pueden pasar a través de las membranas lipídicas de las células endoteliales; estas sustancias son las propias moléculas de agua, losiones sodio y cloruro y la glucosa.
Aunque 1/1.000 de la superficie de los capilares está representada por los espacios intercelulares entre las células endoteliales, la velocidad del movimiento térmico molecular en estos espacios es tan alta que incluso esta pequeña superficie es suficiente para permitir una difusión enorme de agua y sustancias hidrosolubles a través de estos espacios-poros.
EFECTO DEL TAMAÑO MOLECULAR SOBRE EL PASO A TRAVÉS DE LOS POROS
La profundidad de los espacios intercelulares capilares, 6 a 7 nm, es unas 20 veces el diámetro de la molécula de agua, que es la molécula más pequeña que normalmente atraviesa los poros de los capilares.
Sin embargo, los diámetros de las moléculas proteicas plasmáticas son ligeramente mayores que la anchura de los poros. Otras sustancias, como los iones sodio o cloruro, la glucosa y la urea, tienen diámetros intermedios.
Por tanto, la permeabilidad de los poros del capilar para distintas sustancias varía según sus diámetros moleculares.
PERMEABILIDAD RELATIVA DE LOS POROS CAPILARES EN EL MÚSCULO ESQUELÉTICO SEGÚN LOS DISTINTOS TAMAÑOS DE LAS MOLECULAS
En este punto tenemos que hacer una salvedad. Los capilares de los diversos tejidos tienen diferencias extremas en su permeabilidad.
Por ejemplo, las membranas de los sinusoides del capilar hepático son tan permeables que incluso las proteínas plasmáticas atraviesan esas paredes casi tan fácilmente como el agua y otras sustancias.
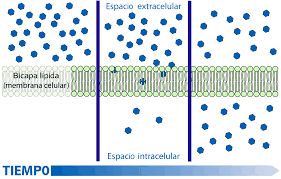
EFECTO DE LA DIFERENCIA DE CONCENTRACIÓN EN LA VELOCIDAD NETA DE DIFUSIÓN A TRAVÉS DE LA MEMBRANA CAPILAR
La velocidad «neta» de difusión de una sustancia a través de cualquier membrana es proporcional a ladiferencia de concentración de la sustancia entre los dos lados de la membrana.
Es decir, cuanto mayor sea la diferencia entre las concentraciones de una sustancia dada en los dos lados de la membrana capilar, mayor será el movimiento neto de la sustancia en una dirección a través de la membrana.
Por ejemplo, la concentración de oxígeno en la sangre capilar es generalmente mayor que en el líquido intersticial.
Por tanto, normalmente se mueven grandes cantidades de oxígeno desde la sangre hacia los tejidos, mientras que, por el contrario, la concentración de dióxido de carbono es mayor en los tejidos que en la sangre, lo que hace que el exceso de dióxido de carbono se mueva hacia la sangre y se transporte lejos de los tejidos.
INTERSTICIO Y LÍQUIDO INTERSTICIAL
Una sexta parte del volumen total del organismo consiste en espacios comprendidos entre las células y que se define colectivamente como intersticio.
El líquido que circula a través de estos espacios se denomina líquido intersticial.
Este intersticio esta constituido por dos tipos de estructuras solidas: 1: Haces de fibras de colágeno y 2: Filamentos de proteoglucano.
Los haces de fibras de colágeno recorren largas distancias dentro del intersticio, son muy fuertes y le proporcionan la mayor parte de la fuerza tensiones a los tejidos.
Por el contrario los filamentos de proteoglucano son moléculas muy finas enrolladas o retorcidas constituidas por el 98% de acido hialuronico y el 2% de proteínas. No pueden observarse en el microscopio óptico y en el electrónico es difícilmente apréciarles.
Su importancia radica en el hecho de que forman filamentos reticulares muy finos, que se describen como bordes en cepillo.
GEL EN EL INTERSTICIO
El líquido intersticial deriva por filtración y difusión de los capilares. Contiene los mismos componentes que el plasma a excepción de que tiene una concentración más baja de proteínas por que estas no pueden atravesar los poros de los capilares.
El líquido intersticial queda atrapado en
El líquido intersticial queda atrapado principalmente en los diminutos espacios que hay entre los filamentos de proteoglucanos. Esta combinación de filamentos de proteoglucano y líquido atrapado dentro de ellos tiene las características de un gel y, por tanto, se conoce como gel tisular.
Debido al gran número de filamentos de proteoglucano, es difícil que el líquido fluya fácilmente a través de este gel tisular. Por el contrario, el líquido se difunde principalmente a través del gel, es decir, se desplaza molécula a molécula de un lugar a otro por un movimiento cinético térmico y no por el gran número de moléculas que se desplazan juntas.
La difusión a través del gel se produce con una rapidez del 95 al 99% de la que se desplaza a través de un líquido libre.
LIQUIDO LIBRE EN EL INTERSTICIO
Aunque casi todo el líquido del intersticio está atrapado dentro del gel tisular, a veces también hay pequeños riachuelos de líquido «libre» y pequeñas vesículas de líquido libre, lo que significa que carece de moléculas de proteoglucano y, por tanto, puede fluir libremente.
Cuando se inyecta un colorante en la sangre circulante, a menudo puede verse fluir a través del intersticio en los riachuelos pequeños que circulan habitualmente por las superficies de las fibras de colágeno o en las superficies de las células.
LA FILTRACIÓN DE LÍQUIDOS A TRAVÉS DE LOS CAPILARES SE ENCUENTRA DETERMINADA POR LAS PRESIONES HIDROSTÁTICA Y COLOIDOSMÓTICA Y POR EL COEFICIENTE DE FILTRACIÓN CAPILAR
La presión hidrostática en los capilares tiende a empujar el líquido y a las sustancias disueltas a través de los poros capilares dentro de los espacios intersticiales. Por el contrario, la presión osmótica provocada por las proteínas plasmáticas (lo que se conoce como presión coloidosmótica) tiende a provocar el movimiento del líquido por ósmosis desde los espacios intersticiales hacia la sangre.
Esta presión osmótica ejercida por las proteínas plasmáticas normalmente previene la pérdida significativa de volumen de líquido desde la sangre hacia los espacios intersticiales.
LAS FUERZAS HIDROSTATICAS Y COLOIDOSMOTICA DETERMINAN EL MOVIMIENTO DE LÍQUIDO A TRAVÉS DE LA MEMBRANA CAPILAR
1. La presión capilar (Pc), que tiende a forzar la salida del líquido a través de la membrana capilar.
2. La presión del líquido intersticial (Pif), que tiende a forzar la entrada del líquido a través de la membrana capilar cuando la Pif es positiva, pero fuerza la salida cuando la Pif es negativa.
3. La presión coloidosmótica del plasma en el capilar (Πp), que tiende a provocar ósmosis de líquido hacia el interior a través de la membrana capilar.
4. La presión coloidosmótica del líquido intersticial (Πif), que tiende a provocar la ósmosis del líquido hacia el exterior a través de la membrana capilar.
Si la suma de estas fuerzas, la presión de filtración neta, es positiva, habrá una filtración neta de líquidos a través de los capilares. Si la suma de las fuerzas de Starling es negativa, habrá una absorción neta de líquido desde los espacios intersticiales hacia los capilares. La presión neta de filtración (PNF) se calcula como:
Por tanto, la velocidad de la filtración de líquidos en el capilar está determinada por:
PRESIÓN HIDROSTÁTICA CAPILAR
Se han usado varios métodos experimentales para estimar la presión hidrostática capilar:
1) canulación directa de los capilares con la micropipeta, que da una presión capilar media de 25 mmHg en algunos tejidos como el músculo esquelético y el aparato digestivo, y
2) determinación funcional
MÉTODO DE LA MICROPIPETA PARA MEDIR LA PRESIÓN CAPILAR
Para medir la presión por canulación se empuja una pipeta de vidrio microscópica directamente en el capilar y la presión se mide por un sistema apropiado con un micromanómetro. Usando este método se ha medido la presión capilar en los capilares de los tejidos expuestos de animales y en los grandes bucles capilares del eponiquio en la base de las uñas en el ser humano.
MÉTODO ISOGRAVIMETRICO PARA LA MEDICIÓN FUNCIONAL INDIRECTA DE LA PRESIÓN CAPILAR
En la figura se muestra un método isogravimétrico para estimar indirectamente la presión capilar. En esta figura se muestra una sección del intestino sujeta por uno de los brazos de una balanza gravimétrica.
La sangre se perfunde a través de los vasos sanguíneos de la pared intestinal. Cuando la presión arterial disminuye, el descenso resultante de la presión capilar permite que la presión osmótica de las proteínas plasmáticas provoque la absorción de líquido hacia el interior de la pared intestinal y hace que el peso del intestino disminuya, lo que inmediatamente consigue el desplazamiento del brazo de la balanza.
PRESIÓN HIDROSTATICA DEL LÍQUIDO INTERSTICIAL
Hay varios métodos que permiten medir la presión hidrostática del líquido intersticial, cada uno de los cuales da valores ligeramente distintos, en función del método utilizado y del tejido en el que se mide la presión.
Los métodos más usados han sido:
1) medida directa de la presión con una micropipeta introducida en los tejidos;
2) determinación de la presión desde cápsulas perforadas implantadas,
y 3) determinación de la presión desde una mecha de algodón insertada en el tejido. Estos distintos métodos proporcionan valores diferentes de la presión hidrostática intersticial, incluso en los mismos tejidos.
PRESIÓN DEL LÍQUIDO INTERSTICIAL EN TEJIDOS FIRMEMENTE ENCAPSULADOS
Algunos tejidos del organismo están rodeados por una carcasa rígida, como la bóveda craneal alrededor del cerebro, la cápsula fibrosa fuerte que rodea el riñón, las vainas fibrosas de los músculos y la esclerótica que rodea el ojo.
En la mayoría de estos tejidos, e independientemente del método usado para la determinación, la presión del líquido intersticial suele ser positiva. No obstante, la presión del líquido intersticial así medida casi invariablemente es aún menor que las presiones ejercidas en el exterior de los tejidos por sus carcasas.
Algunas de estas cavidades y las determinaciones de las presiones son las siguientes:
• Espacio intrapleural: –8 mmHg.
• Espacio sinovial articular: –4 a –6 mmHg.
• Espacio epidural: –4 a –6 mmHg.
RESUMEN: LA PRESIÓN NEGATIVA DEL LÍQUIDO INTERSTICIAL EN EL TEJIDO SUBCUTANEO LAXO SUELE SUBATMOSFERICA
Aunque los distintos métodos mencionados anteriormente obtienen valores discretamente diferentes de la presión del líquido intersticial, la mayoría de los fisiólogos creen que la presión del líquido intersticial en el tejido subcutáneo laxo es, en condiciones normales, algo menor que la atmosférica, con un promedio de –3 mmHg.
LA FUNCIÓN DE BOMBA DEL SISTEMA LINFÁTICO ES LA CAUSA BÁSICA DE LA PRESIÓN NEGATIVA DEL LÍQUIDO INTERSTICIAL
El sistema linfático se comenta más adelante en este mismo capítulo, pero primero tenemos que conocer la función básica que tiene este sistema para determinar la presión del líquido intersticial. El sistema linfático es un sistema «eliminador» que extrae el exceso de líquido, el exceso de moléculas proteicas, los restos celulares y otras sustancias de los espacios tisulares.
LAS PROTEÍNAS PLASMATICAS CREAN LA PRESIÓN COLOIDOSMOTICA
Como las proteínas son los únicos componentes disueltos en el plasma y en el líquido intersticial que no atraviesan fácilmente los poros capilares, son las responsables de las presiones osmóticas a ambos lados de la membrana capilar.
Para distinguir esta presión osmótica que se produce en la membrana celular se denomina presión coloidosmótica o presión oncótica. El término presión osmótica «coloide» se debe a que la solución de proteínas se parece a una solución coloidal, a pesar de que realmente es una solución molecular verdadera.
VALORES NORMALES DE PRESIÓN COLOIDOSMOTICA DEL PLASMA
La presión coloidosmótica del plasma humano normal alcanza un promedio de 28 mmHg, de los que 19 mm se deben a los efectos moleculares de las proteínas disueltas y 9 mm al efecto de Donnan, es decir, a la presión osmótica extra causada por el sodio, el potasio y los demás cationes que las proteínas mantienen en el plasma.
PRESIÓN COLOIDOSMOTICA DEL LÍQUIDO INTERSTICIAL
La cantidad total de proteínas en los 12 l de líquido intersticial del organismo es ligeramente mayor que la cantidad total de proteínas en el propio plasma, pero, como su volumen es cuatro veces mayor que el volumen del plasma, la concentración media de proteínas en el líquido intersticial de la mayoría de los tejidos solo es del 40% de la plasmática, unos 3 g/dl. Cuantitativamente, la presión coloidosmótica media del líquido intersticial para esta concentración de proteínas es de 8 mmHg.
INTERCAMBIO DE VOLUMEN DE LÍQUIDO A TRAVÉS DE LA MEMBRANA CAPILAR
Ahora que ya hemos comentado los distintos factores que afectan al movimiento del líquido a través de la membrana capilar, podemos ponerlos todos juntos y ver cómo el siste- ma capilar mantiene la distribución normal del volumen de líquido entre el plasma y el líquido intersticial.
La presión capilar media en los extremos arteriales de los capilares es 15 a 25 mmHg mayor que en los extremos venosos. Debido a esta diferencia, el líquido «se filtra» fuera de los capilares en los extremos arteriales, pero en los extremos venosos vuelve a ser reabsorbido en los capilares.
ANALISIS DE LAS FUERZAS QUE PROVOCAN LA FILTRACIÓN EN EL EXTREMO ARTERIAL DEL CAPILAR
Las fuerzas medias apropiadas que están operativas en el extremo arterial del capilar que provocan el movimiento a través de la membrana capilar son las siguientes:
ANALISIS DE LA REABSORCIÓN EN EL EXTREMO VENOSO CAPILAR
La presión sanguínea baja en el extremo venoso del capilar cambia el equilibrio de fuerzas a favor de la absorción, de la siguiente forma:
EQUILIBRIO DE STARLING PARA EL INTERCAMBIO CAPILAR
Ernest Starling señaló hace más de un siglo que, en condiciones normales, existe un estado cercano al equilibrio en la mayoría de los capilares, es decir, que la cantidad de líquido que se filtra de los extremos arteriales de los capilares hacia el exterior es casi exactamente igual a la de líquido que vuelve a la circulación mediante absorción. El ligero desequilibrio que se produce explica el líquido que puede volver a la circulación a través de los vasos linfáticos.
COEFICIENTE DE FILTRACIÓN CAPILAR
En el ejemplo anterior, un desequilibrio neto de las fuerzas en las membranas capilares de 0,3 mmHg provoca la filtración neta de líquidos en todo el organismo de 2 ml/min.
Al expresar la velocidad de filtración neta de líquidos según el desequilibrio por cada milímetro de mercurio se encuentra una velocidad de filtración neta de 6,67 ml/min de líquido por minuto por mmHg para todo el organismo.
Este valor recibe el nombre de coeficiente de filtración capilar corporal total.
EFECTO DE LA ALTERACIONES DEL EQUILIBRIO DE FUERZAS EN LA MEMBRANA CAPILAR
Si la presión capilar media aumenta por encima de 17 mmHg aumenta también la fuerza neta que tiende a causar la filtración de líquido en los espacios tisulares. Es decir, un aumento de 20 mmHg de la presión capilar media provoca un incremento de la presión de filtración de 0,3 mmHg a 20,3 mmHg, con lo que se consigue una filtración neta hasta 68 veces mayor que la que ocurre normalmente en los espacios intersticiales.
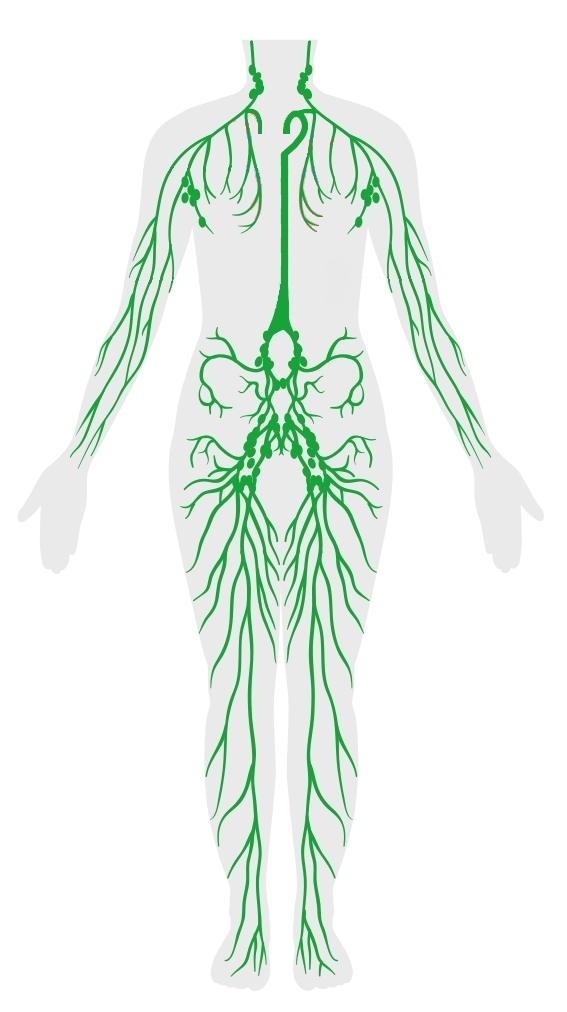
SISTEMA LINFÁTICO
El sistema linfático representa una vía accesoria a través de la cual el líquido puede fluir desde los espacios intersticiales hacia la sangre. Es más, los vasos linfáticos transportan las proteínas y las macropartículas de los espacios tisulares, ya que ninguna de estas podrá ser eliminada por absorción directamente hacia los capilares sanguíneos. Este retorno de las proteínas a la sangre desde los espacios intersticiales es una función esencial sin la cual moriríamos en 24 h.
VASOS LINFATICOS DEL ORGANISMO
Casi todos los tejidos del organismo tienen vasos linfáticos especiales que drenan el exceso de líquido directamente desde los espacios intersticiales. Hay algunas excepciones, como las porciones superficiales de la piel, el sistema nervioso central y el endomisio de músculos y huesos.
Sin embargo, incluso estos tejidos tienen canales intersticiales diminutos que se denominan canales prelinfáticos, a través de los cuales puede fluir el líquido intersticial; este líquido se vacía finalmente en los vasos linfáticos o, en caso del cerebro, en el líquido cefalorraquídeo, y después directamente de vuelta a la sangre.
CAPILARES LINFATICOS TERMINALES Y SU PERMEABILIDAD
La mayoría del líquido que se filtra desde los extremos arteriales de los capilares sanguíneos fluye entre las células y, por último, se reabsorbe de nuevo hacia los extremos venosos de los capilares sanguíneos; pero, como media, aproximadamente la décima parte del líquido entra en los capilares linfáticos y vuelve hacia la sangre a través del sistema linfático y no al contrario, a través de los capilares venosos. La cantidad total de toda esta linfa normalmente es de solo 2-3 l al día.
El líquido que vuelve a la circulación a través de los linfáticos es muy importante porque las sustancias de alto peso molecular que contiene, como las proteínas, no pueden ser absorbidas desde los tejidos de ninguna otra forma, aunque pueden entrar en los capilares linfáticos casi sin impedimentos.
Una característica muy importante del aparato vascular es que todos los vasos sanguíneos sondistensibles. La naturaleza distensible de las arterias las permite acomodarse al gasto pulsátil del corazón y superar las pulsaciones de la presión. Esta capacidad proporciona un flujo de sangre continuo y homogéneo a través de los vasos sanguíneos muy pequeños de los tejidos.
Con diferencia, los vasos más distensibles del cuerpo son las venas, capaces de almacenar 0,5-1 l de sangre extra con incrementos incluso leves de la presión venosa. Por tanto, las venas ejercen de reservorio para almacenar grandes cantidades de sangre extra que puede utilizarse siempre que se requiera en cualquier otro punto de la circulación.
UNIDADES DE DISTENSIBILIDAD VASCULAR
La distensibilidad vascular se expresa como el incremento fraccionado del volumen por cada milímetro de mercurio que aumenta la presión, según la fórmula:
Es decir, si 1 mmHg provoca el aumento de volumen de 1 ml en un vaso que originalmente contenía 10 mm de sangre, la distensibilidad sería de 0,1 por mmHg o del 10% por mmHg.
LAS VENAS SON MUCHO MAS DISTENSIBLES QUE LAS ARTERIAS
Las paredes de las arterias son más gruesas y bastante más fuertes que las de las venas, por lo que, como media, las venas son unas ocho veces más distensibles que las arterias. Es decir, un incremento dado de la presión provoca un incremento de sangre ocho veces mayor en una vena que en una arteria de tamaño comparable.
COMPLIANCIA VASCULAR (O CAPACITANCIA VASCULAR)
En los estudios hemodinámicos es mucho más importante conocer la cantidad total de sangre que se puede almacenar en una porción dada de la circulación por cada milímetro de mercurio que aumente la presión que conocer la distensibilidad de cada vaso en particular. Este valor se conoce comocompliancia o capacitancia del lecho vascular respectivo, es decir:
CURVAS DE VOLUMEN-PRESIÓN DE LAS CIRCULACIONES ARTERIAL Y VENOSA
La curva de volumen-presión es una forma cómoda de expresar la relación presión-volumen en un vaso o en cualquier porción de la circulación.
Las curvas trazadas con líneas continuas en rojo y azul de la figura representan, respectivamente, las curvas de volumen-presión del sistema arterial y sistema venoso sistémico normal, demostrando que cuando el sistema arterial de un adulto normal (con todas sus arterias grandes, pequeñas y arteriolas) se llena con 700 ml de sangre, la presión arterial media es de 100 mmHg, pero la presión cae a cero cuando se llena con solo 400 ml.
EFECTO DE LA ESTIMULACIÓN O DE LA INHIBICIÓN SIMPÁTICA SOBRE LAS RELACIONES VOLUMEN-PRESIÓN EN LOS SISTEMAS ARTERIAL Y VENOSO
Es evidente que el aumento del tono del músculo liso vascular provocado por la estimulación simpática aumenta la presión en cada volumen de arterias o venas, mientras que la inhibición simpática lo disminuye.
Este control de los vasos por los nervios simpáticos es muy importante para disminuir las dimensiones de un segmento de la circulación, transfiriendo la sangre a otros segmentos.
Por ejemplo, el aumento del tono vascular a través de la circulación sistémica puede provocar el desplazamiento de grandes volúmenes de sangre hacia el corazón, lo que constituye uno de los métodos principales que usa el organismo para aumentar rápidamente la función de bomba cardíaca.
COMPLIANCIA DIFERIDA (RELAJACIÓN POR ESTRÉS) DE LOS VASOS
El término «compliancia diferida» se refiere al hecho de que un vaso expuesto a un aumento de volumen primero muestra un gran incremento de la presión, pero progresivamente se va produciendo un estiramiento diferido del músculo liso en la pared de los vasos que permite que la presión vuelva a la normalidad en un período de minutos u horas, como se muestra en la figura, donde la presión se registra en un segmento pequeño de la vena ocluido en ambos extremos.
PULSACIONES DE LA PRESIÓN ARTERIAL
Una oleada de sangre llena las arterias con cada latido cardíaco. Si no fuera por la distensibilidad del sistema arterial, toda esta sangre nueva tendría que fluir a través de los vasos sanguíneos periféricos casi instantáneamente, solo en la sístole cardíaca, y no se produciría flujo durante la diástole.
No obstante, la complacía del árbol arterial reduce las pulsaciones de la presión hasta que prácticamente desaparecen en el momento en que la sangre alcanza los capilares, por lo que el flujo sanguíneo tisular es principalmente continuo con un escaso carácter pulsátil.
Hay dos factores importantes que afectan a la presión de pulso:
1) el volumen sistólico del corazón,
y 2) la compliancia (distensibilidad total) del árbol arterial. Hay un tercer factor, algo menos importante, que es la característica de la eyección del corazón durante la sístole.
PERFILES ANORMALES DE LA PRESIÓN DE PULSO
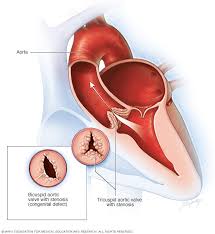
Algunas situaciones fisiopatológicas de la circulación provocan perfiles anormales de la onda de pulsode presión, además de alterar la presión de pulso. Entre ellas, son particularmente importantes la estenosis aórtica, el conducto arterioso permeable y la insuficiencia aórtica
En personas conestenosis valvular aórticael diámetro de apertura de esta válvula está significativamente reducido y la presión de pulso aórtica disminuye también significativamente porque disminuye el flujo sanguíneo que sale por la válvula estenótica.
En personas con conducto arterioso permeable,la mitad o más de la sangre que bombea el ventrículo izquierdo hacia la aorta fluye inmediatamente hacia atrás a través del conducto muy abierto hacia la arteria pulmonar y los vasos sanguíneos pulmonares, con lo que se produce un gran descenso de la presión diastólica antes del siguiente latido cardíaco.
En casos deinsuficiencia aórticaesta válvula está ausente o no se cierra por completo, por lo que después de cada latido la sangre que se acaba de bombear hacia la aorta fluye inmediatamente hacia atrás, hacia el ventrículo izquierdo.
TRANSMISIÓN DE LOS PULSOS DE PRESIÓN HACIA LAS ARTERIAS PERIFÉRICAS
Cuando el corazón expulsa la sangre hacia la aorta durante la sístole, primero se distiende solo la porción proximal de la aorta porque la inercia de la sangre impide el movimiento brusco de la sangre hacia la periferia. No obstante, el aumento de la presión en la aorta proximal supera rápidamente esta inercia y el frente de onda de distensión se va extendiendo a lo largo de la aorta.
Este fenómeno se conoce como transmisión del pulso de presión en las arterias.
LOS PULSOS DE PRESIÓN SE AMORTIGUAN EN LAS ARTERIAS MÁS PEQUEÑAS, ARTERIOLAS Y CAPILARES
En la figura se muestran los cambios típicos del perfil del pulso de presión a medida que se va desplazando hacia los vasos periféricos.
Obsérvese en las tres curvas inferiores que la intensidad de las pulsaciones va siendo progresivamente menor en las arterias más pequeñas, en las arteriolas y, en especial, en los capilares.
De hecho, solo se pueden observar pulsaciones en los capilares cuando la pulsación aórtica es muy grande o cuando las arteriolas están muy dilatadas.
METODOS CLINICOS PARA MEDIR LAS PRESIONES SISTÓLICA Y DIASTÓLICA
No resulta práctico usar registradores de presión que requieran la inserción de la aguja dentro de una arteria para obtener determinaciones sistemáticas de la presión arterial en nuestros pacientes, aunque estos tipos de registradores se utilizan a veces cuando se requieren estudios especiales.
Por el contrario, el médico determina las presiones sistólica y diastólica por medios indirectos, habitualmente por un método de auscultación.
MÉTODO DE AUSCULTACIÓN
Se coloca el estetoscopio sobre la arteria antecubital y se infla un manguito de presión arterial en la parte alta del brazo.
Mientras el manguito comprima el brazo con una presión insuficiente para cerrar la arteria braquial no oiremos el latido de la arteria antecubital con el estetoscopio, pero cuando la presión sea suficientemente elevada para cerrar la arteria durante parte del ciclo de presión arterial se oirá un sonido con cada pulsación.
Estos sonidos se conocen como ruidos de Korotkoff, así llamados por Nikolái Korotkoff, un físico ruso que los describió en 1905.
Según se cree, los ruidos de Korotkoff se deben principalmente al chorro de sangre que atraviesa ese vaso parcialmente ocluido y a las vibraciones de la pared del vaso. El chorro provoca turbulencias del vaso más allá del manguito, con lo que se consigue que las vibraciones se oigan a través del estetoscopio.
Al determinar la presión arterial por este método con auscultación, la presión del manguito primero se eleva por encima de la presión sistólica. Mientras que la presión del manguito sea mayor que la presión sistólica, la arteria braquial se mantiene colapsada hasta que no haya ningún chorro de sangre hacia la parte distal de la arteria en ningún momento del ciclo de presión, por lo que no se oirán ruidos de Korotkoff en la parte distal
A medida que la presión del manguito continúa descendiendo irá cambiando la calidad de los ruidos de Korotkoff, disminuyendo la calidad del ruido y haciéndose más rítmico y duro.
Por último, cuando la presión del manguito desciende casi a los valores de la presión diastólica, los ruidos adquieren súbitamente una calidad amortiguada.
PRESIONES ARTERIALES ANORMALES MEDIDAS POR EL MÉTODO DE AUSCULTACIÓN
En la figura se muestran las presiones arteriales sistólica y diastólica en distintas edades. El incremento progresivo de la presión con la edad es consecuencia de los efectos del envejecimiento sobre los mecanismos de control de la presión sanguínea.
PRESIÓN ARTERIAL MEDIA
La presión arterial media es la media de las presiones arteriales medidas milisegundo a milisegundo en un período de tiempo y no es igual a la media de las presiones sistólica y diastólica, porque, para frecuencias cardíacas normales, se invierte una mayor fracción del ciclo cardíaco en la diástole que en la sístole; así pues, la presión arterial sigue estando más cercana a la presión diastólica que a la presión sistólica durante la mayor parte del ciclo cardíaco.
Por tanto, la presión arterial media está determinada en un 60% por la presión diastólica y en un 40% por la presión sistólica.
LAS VENAS Y SUS FUNCIONES
Las venas proporcionan vías de paso para el flujo de sangre hacia el corazón, pero también realizan otras funciones que son necesarias para el funcionamiento de la circulación.
Especialmente importante es que son capaces de disminuir y aumentar su tamaño, con lo cual pueden almacenar cantidades de sangre pequeñas o grandes y mantener la sangre disponible para cuando la necesite el resto de la circulación.
PRESIONES VENOSAS: PRESIÓN DE LA AURÍCULA DERECHA (PRESIÓN VENOSA CENTRAL) Y PRESIONES VENOSAS PERÍFERICAS
Para entender las distintas funciones de las venas, primero es necesario conocer algo sobre la presión en su interior y sobre los factores que la determinan.
La sangre de todas las venas sistémicas fluye hacia la aurícula derecha del corazón, por lo que la presión del interior de esta cámara se denomina presión venosa central.
La presión en la aurícula derecha está regulada por el equilibrio entre:
1) la capacidad del corazón de bombear la sangre hacia el exterior de la aurícula y el ventrículo derechos hacia los pulmones,
y 2) la tendencia de la sangre a fluir desde las venas periféricas hacia la aurícula derecha.
Algunos de estos factores que aumentan este retorno venoso (y, por tanto, aumentan la presión en la aurícula derecha) son:
1) aumento del volumen de sangre;
2) aumento del tono de los grandes vasos en todo el organismo, con el incremento resultante de las presiones venosas periféricas,
y 3) dilatación de las arteriolas, lo que disminuye la resistencia periférica y permite que el flujo de sangre entre las arterias y las venas sea más rápido.
a presión normal en la aurícula derecha es de 0 mmHg, que es igual a la presión atmosférica en todo el organismo.
Puede aumentar hasta 20 o 30 mmHg en condiciones muy anormales como:
1) insuficiencia cardíaca grave,
o 2) después de una transfusión masiva de sangre, lo que aumenta en gran medida el volumen total de sangre y hace que cantidades excesivas de sangre intenten llegar al corazón desde los vasos periféricos.
RESISTENCIA VENOSA Y PRESIÓN VENOSA PERIFÉRICA
Las venas grandes ejercen tan poca resistencia al flujo sanguíneo cuando están distendidas que la resistencia es casi cero, y prácticamente no tiene importancia.
No obstante, como se muestra en la figura , la mayoría de las venas grandes que entran en el tórax están comprimidas en muchos puntos por los tejidos circundantes, lo que supone un obstáculo al flujo.
Por ejemplo, las venas de los brazos se comprimen en las angulaciones bruscas que forman sobre la primera costilla.
EFECTO DE LA PRESIÓN ELEVADA EN LA AURÍCULA DERECHA SOBRE LA PRESIÓN VENOSA PERIFÉRICA
Cuando la presión en la aurícula derecha aumenta por encima de su valor normal de 0 mmHg, la sangre comienza a volver a las venas grandes. Este retorno de la sangre aumenta el tamaño de estas últimas e incluso los puntos de colapso se abren cuando la presión en la aurícula derecha aumenta por encima de +4 a +6 mmHg.
EFECTO DE LA PRESIÓN ABDOMINAL SOBRE LAS PRESIONES VENOSAS EN LAS PIERNAS
La presión de la cavidad abdominal de una persona en decúbito normalmente alcanza una media de +6 mmHg, pero puede aumentar hasta +15 o +30 mmHg como consecuencia del embarazo, de tumores grandes, de obesidad abdominal o de la presencia de líquido excesivo (lo que se conoce como «ascitis») en la cavidad abdominal.
EFECTO DE LA PRESIÓN GRAVITACIONAL SOBRE LA PRESIÓN VENOSA
En cualquier organismo de agua que esté expuesto al aire, la presión en la superficie del agua es igual a la presión atmosférica, pero aumenta 1 mmHg por cada 13,6 mm de distancia por debajo de la superficie. Esta presión es consecuencia del peso del agua y, por tanto, se denomina presión gravitacional o hidrostática.
La presión gravitacional también se produce en el aparato vascular del ser humano por el peso de la sangre en las venas.
En las venas de los brazos la presión a nivel de la costilla superior es de +6 mmHg por la compresión de la vena subclavia cuando pasa por encima de ella, pero la presión gravitacional al bajar por el brazo está determinada por la distancia que hay por debajo de esta costilla, es decir, si la diferencia gravitacional entre el nivel de la costilla y la mano es de +29 mmHg, esta presión gravitacional se suma a los +6 mmHg de presión provocados por la compresión de la vena cuando atraviesa la costilla, con lo que obtenemos un total de +35 mmHg de presión en las venas de la mano.
Las venas del cuello de una persona que esté de pie se colapsan casi por completo en todo su recorrido hasta el cráneo, por la presión atmosférica que hay fuera del cuello. Este colapso hace que la presión en estas venas se mantenga en cero durante todo su trayecto.
EFECTO DEL FACTOR GRAVITACIONAL SOBRE LA PRESIÓN ARTERIAL Y OTRAS PRESIONES
El factor gravitacional también afecta a las presiones de las arterias periféricas y los capilares.
Por ejemplo, una persona en bipedestación que tiene una presión arterial media de 100 mmHg a la altura del corazón tiene una presión arterial en los pies en torno a 190 mmHg. Por tanto, cuando se afirma que la presión arterial es de 100 mmHg, se está diciendo que esta es la presión a nivel gravitacional del corazón, pero no necesariamente en otra parte del territorio arterial.
VALVULAS VENOSAS Y BOMBA VENOSA: EFECTO SOBRE LA PRESIÓN VENOSA
Si no hubiera válvulas en las venas el efecto de la presión gravitacional haría que la presión venosa de los pies fuera siempre de +90 mmHg en un adulto en bipedestación.
No obstante, cada vez que se mueven las piernas, se tensan los músculos y se comprimen las venas de los músculos y de los territorios adyacentes, lo que empuja la sangre fuera de ese territorio venoso. Sin embargo, las válvulas de las venas, están distribuidas de tal forma que la dirección del flujo sanguíneo venoso solo puede ir hacia el corazón.
LA INCOMPETENCIA DE LA VALVULA VENOSA PROVOCA LAS VENAS VARICOSAS
Las válvulas del sistema venoso pueden volverse «incompetentes» o incluso llegan a destruirse, con frecuencia cuando las venas han sido objeto de un sobreestiramiento debido a una presión venosa excesiva que se ha mantenido durante semanas o meses, como sucede en el embarazo o cuando se está de pie la mayoría del tiempo.
El estiramiento de las venas aumenta su superficie transversal, pero las valvas de las válvulas no aumentan de tamaño, por lo que ya no se pueden cerrar completamente.
Cuando seproduce esta falta de cierre completo, la presión de las venas de las piernas aumenta en gran medida por el fracaso de la bomba venosa, lo que además aumenta el tamaño de las venas y, finalmente, destruye completamente todas las válvulas.
ESTIMACIÓN CLÍNICA DE LA PRESIÓN VENOSA
La presión venosa puede estimarse observando simplemente el grado de distensión de las venas periféricas, en especial de las venas del cuello.
Por ejemplo, en sedestación las venas del cuello nunca deben estar distendidas en una persona tranquila y en reposo, pero cuando la presión de la aurícula derecha aumenta hasta +10 mmHg, las venas de la parte inferior del cuello comienzan a protruir y todas las venas del cuello están distendidas cuando la presión auricular es de +15 mmHg.
DETERMINACIÓN DIRECTA DE LA PRESIÓN VENOSA Y DE LA PRESIÓN DE LA AURÍCULA DERECHA
La presión venosa también se puede medir si, con cuidado, se introduce una aguja directamente en la vena y se conecta a un registrador de presión.
El único medio que permite medir con exactitud la presión en la aurícula derecha consiste en insertar un catéter a través de las venas periféricas hasta esa cámara.
NIVEL DE REFERENCIA DE LA PRESIÓN PARA MEDIR LA PRESIÓN VENOSA Y OTRAS PRESIONES CIRCULATORIAS
Hasta este momento hemos hablado de que la presión medida en la aurícula derecha es de 0 mmHg y que la presión arterial es de 100 mmHg, pero no hemos hablado del nivel gravitacional del sistema circulatorio al cual se refiere esta presión.
Hay un punto del sistema circulatorio en el que los factores de presión gravitacional provocados por los cambios de posición del cuerpo de una persona sana no afectan a la determinación de la presión en más de 1-2 mmHg en una medición realizada en la válvula tricúspide o cerca de ella, como se ve en el cruce de ejes.
FUNCIÓN DE RESERVORIO DE SANGRE EN LAS VENAS
Más del 60% de toda la sangre venosa del sistema circulatorio suele encontrarse en las venas.
Por este motivo, y porque las venas son tan distensibles, se dice que el sistema venoso actúa como un reservorio sanguíneo en la circulación.
Cuando la sangre sale del organismo y la presión arterial comienza a caer, se activan señales nerviosas desde los senos carotídeos y otras zonas de la circulación sensibles a la presión.
RESERVORIOS SANGUÍNEOS ESPECÍFICOS
Algunas porciones del sistema circulatorio también son tan extensas o distensibles que se conocen como «reservorios sanguíneos específicos».
Estos reservorios incluyen:
1) el bazo, cuyo tamaño a veces disminuye tanto como para liberar hasta 100 ml de sangre hacia otras áreas de la circulación;
2) el hígado, cuyos senos liberan varios cientos de mililitros de sangre hacia el resto de la circulación;
3) las venas abdominales grandes, que contribuyen hasta con 300 ml,
y 4) los plexos venosos situados bajo la piel, que pueden contribuir también con varios cientos de mililitros.
EL BAZO COMO RESERVORIO PARA ALMACENAR ERITROCITOS
En la figura se muestra que el bazo tiene dos áreas independientes para almacenar la sangre: lossenos venosos y la pulpa.
Los senos pueden ingurgitarse igual que cualquier otra parte del sistema venoso y almacenar sangre total.
En la pulpa del bazo los capilares son tan permeables que la sangre total, incluidos los eritrocitos, rezuma a través de las paredes de los capilares hacia la malla trabecular, formando la pulpa roja.
Los eritrocitos quedan atrapados por las trabéculas, mientras que el plasma fluye hacia los senos venosos y después hacia la circulación general.
En consecuencia, la pulpa roja del bazo es un reservorio especial que contiene grandes cantidades de eritrocitos concentrados que pueden expulsarse a la circulación general siempre que el sistema nervioso simpático se excite y provoque que el bazo y sus vasos se contraigan.
Se pueden liberar hasta 50 ml de eritrocitos concentrados hacia la circulación, elevando el hematocrito en un 1-2%.
FUNCIÓN DE LIMPIEZA DE LA SANGRE EN EL BAZO: ELIMINACIÓN DE LAS CÉLULAS VIEJAS
Las células sanguíneas que atraviesan la pulpa esplénica antes de entrar en los senos son cuidadosamente exprimidas, por lo que se puede esperar que los eritrocitos frágiles no superen este traumatismo.
Por tal motivo, muchos de los eritrocitos destruidos en el organismo encuentran su destino final en el bazo. Después de la rotura de las células la hemoglobina liberada y el estroma celular son digeridos por las células reticuloendoteliales del bazo y los productos de la digestión son reutilizados en su mayoría en el organismo como nutrientes, a menudo para elaborar células sanguíneas nuevas.
La función de la circulación consiste en atender las necesidades del organismo: transportar nutrientes hacia los tejidos del organismo, transportar los productos de desecho, transportar las hormonas de una parte del organismo a otra y, en general, mantener un entorno apropiado en todos los líquidos tisulares del organismo para lograr la supervivencia y una funcionalidad óptima de las células.
CARACTERÍSTICAS FÍSICAS DE LA CIRCULACIÓN
La circulación, está divida en circulación sistémica y circulación pulmonar. Como la circulación sistémica aporta el flujo sanguíneo a todos los tejidos del organismo excepto los pulmones, también se conoce como circulación mayor o circulación periférica.
COMPONENTES FUNCIONALES DE LA CIRCULACIÓN
La función de las arteriasconsiste en transportar la sangre con una presión alta hacia los tejidos, motivo por el cual las arterias tienen unas paredes vasculares fuertes y unos flujos sanguíneos importantes con una velocidad alta.
Las arteriolasson las últimas ramas pequeñas del sistema arterial y actúan controlando los conductosa través de los cuales se libera la sangre en los capilares. Las arteriolas tienen paredes musculares fuertes que pueden cerrarlas por completo o que pueden, al relajarse, dilatar los vasos varias veces, con lo que pueden alterar mucho el flujo sanguíneo en cada lecho tisular en respuesta a sus necesidades.
La función de los capilaresconsiste en el intercambio de líquidos, nutrientes, electrólitos, hormonas y otras sustancias en la sangre y en el líquido intersticial. Para cumplir esta función, las paredes del capilar son finas y tienen muchos poros capilares diminutos, que son permeables al agua y a otras moléculas pequeñas.
Las vénulasrecogen la sangre de los capilares y después se reúnen gradualmente formando venas de tamaño progresivamente mayor.
Las venasfuncionan como conductos para el transporte de sangre que vuelve desde las vénulas al corazón; igualmente importante es que sirven como una reserva importante de sangre extra. Como la presión del sistema venoso es muy baja, las paredes de las venas son finas.
VOLÚMENES DE SANGRE EN LOS DISTINTOS COMPONENTES DE LA CIRCULACIÓN
Aproximadamente el 84% de todo el volumen de sangre del organismo se encuentra en la circulación sistémica y el 16% en el corazón y los pulmones. Del 84% que está en la circulación sistémica, aproximadamente el 64% está en las venas, el 13% en las arterias y el 7% en las arteriolas y capilares sistémicos. El corazón contiene el 7% de la sangre, y los vasos pulmonares, el 9%.
SUPERFICIES TRANSVERSALES Y VELOCIDADES DEL FLUJO SANGUÍNEO
Si todos los vasos sistémicos de cada tipo se pusieran uno al lado del otro, la superficie transversal total aproximada para un ser humano medio sería la siguiente:
Obsérvese en particular que la superficie transversal de las venas es mucho mayor que la de las arterias, con una media cuatro veces mayor en las primeras.
Esta diferencia explica la gran capacidad de reserva de sangre en el sistema venoso comparado con el sistema arterial.Como debe pasar el mismo volumen de flujo sanguíneo (F) a través de cada segmento de la circulación en cada minuto, la velocidad del flujo sanguíneo (v) es inversamente proporcional a la superficie transversal vascular (A).
PRESIONES EN LAS DISTINTAS PORCIONES DE LA CIRCULACIÓN
Como el corazón bombea la sangre continuamente hacia la aorta, la presión media en este vaso es alta, con una media en torno a los 100 mmHg. Además, como el bombeo cardíaco es pulsátil, la presión arterial alterna entre una presión sistólica de 120 mmHg y una diastólica de 80 mmHg.
A medida que el flujo sanguíneo atraviesa la circulación sistémica, la presión media va cayendo progresivamente hasta llegar casi a 0 mmHg en el momento en el que alcanza la terminación de las venas cava superior e inferior, donde se vacía en la aurícula derecha del corazón.
La presión de los capilares sistémicos oscila desde 35 mmHg cerca de los extremos arteriolares hasta tan solo 10 mmHg cerca de los extremos venosos, pero la presión media «funcional» en la mayoría de los lechos vasculares es de 17 mmHg, aproximadamente, una presión suficientemente baja que permite pequeñas fugas de plasma a través de los poros diminutos de las paredes capilares, aunque los nutrientespueden difundir fácilmente a través de los mismos poros hacia las células de los tejidos externos.
PRINCIPIOS BÁSICOS DE LA FUNCIÓN CIRCULATORIA
Aunque la función circulatoria es muy compleja, hay tres principios básicos que subyacen en todas las funciones del sistema.
1. El flujo sanguíneo en la mayoría de los tejidos está controlado según la necesidad tisular. Cuando los tejidos son activos necesitan un aporte mucho mayor de nutrientes y, por tanto, un flujo sanguíneo mucho mayor que en reposo, en ocasiones hasta 20 o 30 veces el nivel de reposo, a pesar de que el corazón normalmente no puede aumentar su gasto cardíaco en más de 4-7 veces su gasto cardíaco por encima del nivel en reposo.
2. El gasto cardíaco es la suma de todos los flujos locales de los tejidos. Cuando el flujo sanguíneo atraviesa un tejido, inmediatamente vuelve al corazón a través de las venas y el corazón responde automáticamente a este aumento del flujo aferente de sangre bombeándolo inmediatamente hacia las arterias.
3. La regulación de la presión arterial es generalmente independiente del control del flujo sanguíneo local o del control del gasto cardíaco. El sistema circulatorio está dotado de un extenso sistema de control de la presión arterial.
INTERRELACIONES ENTRE LA PRESIÓN, EL FLUJO Y LA RESISTENCIA
El flujo sanguíneo que atraviesa un vaso sanguíneo está determinado por dos factores:
1) diferencia de presión de la sangre entre los dos extremos de un vaso, también denominado «gradiente de presión» en el vaso, que empuja la sangre a través del vaso,
y 2) los impedimentos que el flujo sanguíneo encuentra en el vaso, que se conoce como resistencia vascular.
El flujo a través del vaso se puede calcular con la fórmula siguiente, que se conoce como ley de Ohm:
Debido a la gran importancia que tiene esta fórmula, también deberá familiarizarse con sus demás formas algebraicas:
FLUJO SANGUÍNEO
El flujo sanguíneo es, sencillamente, la cantidad de sangre que atraviesa un punto dado de la circulación en un período de tiempo determinado. Normalmente se expresa en mililitros por minuto o litros por minuto, pero puede expresarse en mililitros por segundo o en cualquier otra unidad del flujo y de tiempo.
El flujo sanguíneo global de toda la circulación de un adulto en reposo es de unos 5.000 ml/min, cantidad que se considera igual al gasto cardíaco porque es la cantidad de sangre que bombea el corazón en la aorta en cada minuto.
METODOS DE MEDICIÓN DEL FLUJO SANGUÍNEO
Se pueden introducir muchos dispositivos mecánicos y electromecánicos en serie dentro de un vaso sanguíneo o bien aplicarse en el exterior de la pared del vaso para medir el flujo. Estos dispositivos se denominan flujómetros.
FLUJÓMETRO ELECTROMAGNÉTICO
Un dispositivo que permite medir experimentalmente el flujo sanguíneo sin abrir el vaso es el flujómetro electromagnético.
La generación de la fuerza electromotriz (voltaje eléctrico) de un cable que se mueve rápidamente atravesando un campo magnético. Este es el principio de producción de electricidad en un generador eléctrico.
FLUJÓMETRO ULTRASÓNICO DE DOPPLER
Otro tipo de flujómetro que puede aplicarse al exterior del vaso y que tiene las mismas ventajas que el flujómetro electromagnético es el flujómetro ultrasónico Doppler, que se muestra en la figura. Se monta un cristal piezoeléctrico diminuto en el extremo de la pared del dispositivo.
Cuando este cristal recibe la energía de un aparato eléctrico apropiado transmite una frecuencia de varios cientos de miles de ciclos por segundo distalmente sobre la sangre circulante.
FLUJO DE SANGRE LAMINAR EN LOS VASOS
Cuando el flujo sanguíneo se mantiene en equilibrio a través de un vaso sanguíneo largo y liso, el flujo se produce de forma aerodinámica, manteniéndose cada capa de sangre a la misma distancia de la pared del vaso. Además, la porción de sangre más central se mantiene en el centro del vaso.
Este tipo de flujo se conoce como flujo laminar o flujo aerodinámico y es el contrario del flujo turbulento, que es el flujo sanguíneo que transcurre en todas las direcciones del vaso y se mezcla continuamente en su interior.
PERFIL DE VELOCIDAD PARABÓLICA DURANTE EL FLUJO LAMINAR
Cuando se produce el flujo laminar, la velocidad del flujo en el centro del vaso es bastante mayor que la velocidad cerca de los bordes exteriores
FLUJO DE SANGRE TURBULENTO EN ALGUNAS SITUACIONES
Cuando la velocidad del flujo sanguíneo es demasiado grande, cuando atraviesa una obstrucción en un vaso, hace un giro brusco o pasa sobre una superficie rugosa, el flujo puede volverse turbulento o desordenado en lugar de aerodinámico.
El flujo turbulento significa que el flujo sanguíneo atraviesa el vaso en dirección transversal y también longitudinal, formando espirales que se denominan corrientes en torbellino, similares a los remolinos que se ven con frecuencia en un río que fluye rápidamente en un punto de obstrucción.
El flujo turbulento tiende a aumentar en proporción directa a la velocidad del flujo sanguíneo, al diámetro del vaso sanguíneo y a la densidad de la sangre y es inversamente proporcional a la viscosidad de la sangre, de acuerdo a la ecuación siguiente:
UNIDADES ESTÁNDAR DE PRESIÓN
La presión sanguínea se mide casi siempre en milímetros de mercurio (mmHg) porque el manómetro de mercurio se ha usado como patrón de referencia para medir la presión desde su invención en 1846 por Poiseuille.
En realidad, la presión arterial mide la fuerza ejercida por la sangre contra una unidad de superficie de la pared del vaso. Cuando se dice que la pared de un vaso es de 50 mmHg, quiere decirse que la fuerza ejercida es suficiente para empujar una columna de mercurio contra la gravedad hasta una altura de 50 mm. Si la presión es de 100 mmHg, empujará la columna de mercurio hasta los 100 mm.
MÉTODOS DE ALTA FIDELIDAD PARA MEDIR LA PRESIÓN SANGUÍNEA
El mercurio del manómetro tiene tal inercia que no es capaz de subir y bajar con rapidez, por lo que este aparato, que es excelente para registrar presiones en equilibrio, no puede responder a los cambios de presión que se producen más deprisa que un ciclo cada 2-3 s. Siempre que se desee registrar rápidamente los cambios de presión es necesario utilizar otro tipo de registrador de presión.
UNIDADES DE RESISTENCIA
La resistencia es el impedimento al flujo sanguíneo en un vaso, pero no se puede medir por medios directos. Por el contrario, la resistencia debe calcularse a partir de las determinaciones del flujo sanguíneo y de la diferencia de presión entre dos puntos del vaso.
Si la diferencia de presión entre los dos puntos es de 1 mmHg y el flujo es de 1 ml/s, se dice que la resistencia es de una unidad de resistencia periférica, abreviada habitualmente como PRU.
EXPRESIÓN DE LA RESISTENCIA EN UNIDADES CGS
En ocasiones se usa una unidad física básica en CGS (centímetros, gramos, segundos) para expresar la resistencia. Esta unidad es la dina · s/cm5 . La resistencia en esas unidades puede calcularse mediante la fórmula siguiente:
RESISTENCIA VASCULAR PERIFÉRICA TOTAL Y RESISTENCIA VASCULAR PULMONAR TOTAL
La velocidad del flujo sanguíneo a través de todo el sistema circulatorio es igual a la velocidad de la sangre que bombea el corazón, es decir, es igual al gasto cardíaco. En un ser humano adulto es aproximadamente igual a 100 ml/s. La diferencia de presión entre las arterias sistémicas y las venas sistémicas es de unos 100 mmHg. Por tanto, la resistencia de toda la circulación sistémica, que se denomina resistencia periférica total, es de 100/100 o 1 PRU.
LA CONDUCTANCIA DE LA SANGRE EN UN VASO ES INVERSA A LA RESISTENCIA
La conductancia es la medición del flujo sanguíneo a través de un vaso para dar una diferencia de presión dada. Esta medida se expresa en milímetros por segundo por milímetro de mercurio de presión, pero también se puede expresar en litros por segundo por milímetro de mercurio o en cualquier otra unidad del flujo sanguíneo y presión.
Es evidente que la conductancia es el recíproco exacto de la resistencia según la ecuación:
CAMBIOS PEQUEÑOS EN EL DIÁMETRO DE UN VASO CAMBIAN MUCHO LA CONDUCTANCIA
Pequeños cambios en el diámetro de un vaso provocan cambios enormes en su capacidad de conducir la sangre cuando el flujo sanguíneo es aerodinámico. Este fenómeno se ilustra en el experimento de la figura,en la que vemos tres vasos con diámetros relativos de 1, 2 y 4 pero con la misma diferencia de presión de 100 mmHg entre los dos extremos del vaso.
LEY DE POISEUILLE
Al integrar las velocidades de todos los anillos concéntricos de la sangre en movimiento y multiplicarlos por las superficies de los anillos se puede obtener la fórmula siguiente, que representa la ley de Poiseuille:
IMPORTANCIA DE LA LEY DE LA CUARTA POTENCIA DEL DIÁMETRO DEL VASO PARA DETERMINAR LA RESISTENCIA ARTERIOLAR
En la circulación sistémica, aproximadamente dos tercios de toda la resistencia sistémica al flujo sanguíneo se debe a la resistencia arteriolar en las pequeñas arteriolas. Los diámetros internos de las arteriolas varían desde tan solo 4 μm hasta 25, aunque sus fuertes paredes vasculares permiten cambios enormes de los diámetros internos, a menudo hasta en cuatro veces.
RESISTENCIA AL FLUJO SANGUÍNEO EN CIRCUITOS VASCULARES EN SERIE Y EN PARALELO
La sangre que bombea el corazón fluye desde la parte de presión alta de la circulación sistémica (es decir, la aorta) hacia el lado de baja presión (es decir, la vena cava) a través de muchos miles de vasos sanguíneos dispuestos en serie y en paralelo.
Las arterias, arteriolas, capilares, vénulas y venas se disponen colectivamente en serie. Cuando esto sucede, el flujo de cada vaso sanguíneo es el mismo y la resistencia total al flujo sanguíneo (Rtotal) es igual a la suma de la resistencia de cada vaso:
EFECTO DEL HEMATOCRITO Y DE LA VISCOSIDAD DE LA SANGRE SOBRE LA RESISTENCIA VASCULAR Y EL FLUJO SANGUÍNEO
Otro de los factores importantes de la ley de Poiseuille es la viscosidad de la sangre. Cuanto mayor sea la viscosidad, menor será el flujo en un vaso si todos los demás factores se mantienen constantes. Además, la viscosidad de la sangre normal es tres veces mayor que la del agua.
HEMATOCRITO: PROPORCIÓN DE SANGRE COMPUESTA POR ERITROCITOS
Si una persona tiene un hematocrito de 40 significa que el 40% del volumen sanguíneo está formado por las células y el resto es plasma. El hematocrito de un hombre adulto alcanza un promedio de 42, mientras que en las mujeres es de 38. Estos valores son muy variables, dependiendo de si la persona tiene anemia, del grado de actividad corporal y de la altitud en la que reside la persona.
EL AUMENTO DEL HEMATOCRITO INCREMENTA MUCHO LA VISCOSIDAD DE LA SANGRE
La viscosidad de la sangre aumenta drásticamente a medida que lo hace el hematocrito, como se ve en la figura. La viscosidad de la sangre total con un hematocrito normal es de 3 a 4, lo que significa que se necesita tres veces más presión para obligar a la sangre total a atravesar un vaso que si fuera agua.
Cuando el hematocrito aumenta hasta 60 o 70, como sucede en personas con policitemia, la viscosidad de la sangre puede ser hasta 10 veces mayor que la del agua y su flujo a través de los vasos sanguíneos se retrasa mucho.
Después de que los alvéolos se hayan ventilado con aire limpio, la siguiente fase de la respiración es ladifusión del oxígeno (O2) desde los alvéolos hacia la sangre pulmonar y la difusión del dióxido de carbono (CO2) en la dirección opuesta, desde la sangre a los alvéolos. El proceso de difusión es simplemente el movimiento aleatorio de moléculas en todas las direcciones a través de la membrana respiratoria y los líquidos adyacentes.
BASE MOLECULAR DE LA DIFUSIÓN GASEOSA
Todos los gases importantes en fisiología respiratoria son moléculas simples que se mueven libremente entre sí por «difusión». Esto también se aplica a los gases que están disueltos en los líquidos y en los tejidos del cuerpo.
Para que se produzca la difusión debe haber una fuente de energía. Esta fuente procede del movimiento cinético de las propias partículas. Excepto a la temperatura del cero absoluto, todas las moléculas de toda la materia están experimentando movimiento de manera continua.
DIFUSIÓN NETA DE UN GAS EN UNA DIRECCIÓN: EFECTO DE UN GRADIENTE DE CONCENTRACIÓN
Si una cámara de gas o una solución tiene una concentración elevada de un gas particular en un extremo de la cámara y una concentración baja en el otro extremo, se producirá difusión neta del gas desde la zona de concentración elevada hacia la zona de concentración baja.
La razón es evidente: hay muchas más moléculas en el extremo A de la cámara para difundir hacia el extremo B que moléculas para difundir en la dirección opuesta. Por tanto, las velocidades de difusión en cada una de las dos direcciones son diferentes proporcionalmente.
PRESIONES GASEOSAS EN UNA MEZCLA DE GASES: PRESIONES PARCIALES DE GASES INDIVIDUALES
La presión está producida por múltiples impactos de partículas en movimiento contra una superficie. Por tanto, la presión de un gas que actúa sobre las superficies de las vías aéreas y de los alvéolos es proporcional a la suma de las fuerzas de los impactos de todas las moléculas de ese gas que chocan contra la superficie en cualquier momento dado. Esto significa que la presión es directamente proporcional a la concentración de las moléculas del gas.
El aire, que tiene una composición aproximada del 79% de nitrógeno y el 21% de oxígeno. La presión total de esta mezcla al nivel del mar es en promedio de 760 mmHg. A partir de la descripción previa de la base molecular de la presión es evidente que cada uno de los gases contribuye a la presión total en proporción directa a su concentración.
Por tanto, el 79% de los 760 mmHg está producido por el nitrógeno (600 mmHg) y el 21% por el O2 (160 mmHg). Así, la «presión parcial» del nitrógeno en la mezcla es de 600 mmHg y la «presión parcial» del O2 es de 160 mmHg; la presión totales de 760 mmHg, la suma de las presiones parciales individuales. Las presiones parciales de los gases individuales en una mezcla se señalan por los símbolos Po2, Pco2, Pn2, Phe, etc.
PRESIONES DE GASES DISUELTOS EN AGUA Y TEJIDOS
Los gases disueltos en agua o en los tejidos corporales también ejercen una presión, porque las moléculas de gas disuelto se mueven de manera aleatoria y tienen energía cinética.
Además, cuando el gas disuelto en el líquido entra en contacto con una superficie, como la membrana de una célula, ejerce su propia presión parcial de la misma manera que un gas en la fase gaseosa. Las presiones parciales de diferentes gases disueltos se denominan de la misma manera que las presiones parciales en estado gaseoso, es decir, Po2, Pco2, Pn2, Phe, etc.
FACTORES QUE DETERMINAN LA PRESIÓN PARCIAL DE UN GAS DISUELTO EN UN LÍQUIDO
La presión parcial de un gas en una solución está determinada no solo por su concentración, sino también por el coeficiente de solubilidad del gas. Es decir, algunos tipos de moléculas, especialmente el CO2, son atraídas física o químicamente por las moléculas de agua, mientras que otros tipos de moléculas son repelidas.
Cuando las moléculas son atraídas se pueden disolver muchas más sin generar un exceso de presión parcial en el interior de la solución. Por el contrario, en el caso de moléculas que son repelidas se generará una presión parcial elevada con menos moléculas disueltas. Estas relaciones se expresan mediante la fórmula siguiente, que es la ley de Henry:
DIFUSIÓN DE GASES ENTRE LA FASE GASEOSA DE LOS ALVÉOLOS Y LA FASE DISUELTA DE LA SANGRE PULMONAR
La presión parcial de cada uno de los gases en la mezcla de gas respiratorio alveolar tiende a hacer que las moléculas de ese gas se disuelvan en la sangre de los capilares alveolares.
Por el contrario, las moléculas del mismo gas que ya están disueltas en la sangre están rebotando de manera aleatoria en el líquido de la sangre, y algunas de estas moléculas que rebotan escapan de nuevo hacia los alvéolos. La velocidad a la que escapan es directamente proporcional a su presión parcial en la sangre.
PRESIÓN DE VAPOR DE AGUA
Cuando se inhala aire no humidificado hacia las vías aéreas, el agua se evapora inmediatamente desde las superficies de estas vías aéreas y humidifica el aire.
Esto se debe al hecho de que las moléculas de agua, al igual que las moléculas de los diferentes gases disueltos, están escapando continuamente de la superficie del agua hacia la fase gaseosa.
La presión parcial que ejercen las moléculas de agua para escapar a través de la superficie se denomina la presión de vapor del agua. A la temperatura corporal normal, 37 °C, esta presión de vapor es de 47 mmHg. Por tanto, una vez que la mezcla de gases se ha humidificado totalmente (es decir, una vez que está en «equilibrio» con el agua), la presión parcial del vapor de agua en la mezcla de gases es de 47 mmHg. Esta presión parcial, al igual que las demás presiones parciales, se denomina Ph2o.
CUANTIFICACIÓN DE LA VELOCIDAD NETA DE DIFUSIÓN EN LÍQUIDOS
Además de la diferencia de presión, hay diversos factores que afectan a la velocidad de difusión del gas en un líquido. Estos factores son:
1) la solubilidad del gas en el líquido.
2) el área transversal del líquido.
3) la distancia a través de la cual debe difundir el gas.
4) el peso molecular del gas.
y 5) la temperatura del líquido.
Todos estos factores se pueden expresar en una única fórmula, que es la siguiente:
HUMIDIFICACIÓN DEL AIRE EN LAS VÍAS AÉREAS
La presión parcial de vapor de agua a una temperatura corporal normal de 37 °C es de 47 mmHg, que es, por tanto, la presión parcial de vapor de agua del aire alveolar. Como la presión total en los alvéolos no puede aumentar por encima de la presión atmosférica (760 mmHg a nivel del mar), este vapor de agua simplemente diluye todos los demás gases que están en el aire inspirado.
La humidificación del aire diluye la presión parcial de oxígeno al nivel del mar desde un promedio de 159 mmHg en el aire atmosférico a 149 mmHg en el aire humidificado, y diluye la presión parcial de nitrógeno desde 597 a 563 mmHg.
EL AIRE ALVEOLAR SE RENUEVA LENTAMENTE POR EL AIRE ATMOSFÉRICO
En promedio la capacidad residual funcional de los pulmones (el volumen de aire que queda en los pulmones al final de una espiración normal) en un hombre mide aproximadamente 2.300 ml.
Sin embargo, solo 350 ml de aire nuevo entran en los alvéolos en cada inspiración normal y se espira esta misma cantidad de aire alveolar. Por tanto, el volumen de aire alveolar que es sustituido por aire atmosférico nuevo en cada respiración es de solo 1/7 del total, de modo que son necesarias múltiples inspiraciones para intercambiar la mayor parte del aire alveolar.
CONCENTRACIÓN Y PRESIÓN PARCIAL DE OXÍGENO EN LOS ALVÉOLOS
El oxígeno se absorbe continuamente desde los alvéolos hacia la sangre de los pulmones, y continuamente se respira O2 nuevo hacia los alvéolos desde la atmósfera.
Cuanto más rápidamente se absorba el O2, menor será su concentración en los alvéolos; por el contrario, cuanto más rápidamente se inhale nuevo O2 hacia los alvéolos desde la atmósfera, mayor será su concentración. Por tanto, la concentración de O2 en los alvéolos, y también su presión parcial, está controlada por:
1) la velocidad de absorción de O2 hacia la sangre.
y 2) la velocidad de entrada de O2 nuevo a los pulmones por el proceso ventilatorio.
CONCENTRACIÓN Y PRESIÓN PARCIAL DE CO2 EN LOS ALVÉOLOS
El dióxido de carbono se forma continuamente en el cuerpo y después se transporta por la sangre hacia los alvéolos; se elimina continuamente de los alvéolos por la ventilación.
La figura 40-5 muestra los efectos sobre la presión parcial de CO2 (Pco2) alveolar tanto de la ventilación alveolar como de dos velocidades de excreción de CO2, 200 y 800 ml/min.
Una curva representa una velocidad normal de excreción de CO2 de 200 ml/min. A la velocidad normal de ventilación alveolar de 4,2 l/min, el punto operativo para la Pco2 alveolar está en el punto A de la figura 40-5 (es decir, 40 mmHg).
UNIDAD RESPIRATORIA
La unidad respiratoria (también denominada «lobulillo respiratorio»), está formada por un bronquíolo respiratorio, los conductos alveolares, los atrios y los alvéolos.
Hay aproximadamente 300 millones de alvéolos en los dos pulmones, y cada alvéolo tiene un diámetro medio de aproximadamente 0,2 mm. Las paredes alveolares son muy delgadas y entre los alvéolos hay una red casi sólida de capilares interconectados. De hecho, debido a lo extenso del plexo capilar, se ha descrito que el flujo de sangre en la pared alveolar es una «lámina» de sangre que fluye.
MEMBRANA RESPIRATORIA
La membrana respiratoria esta formada por:
1. Una capa de líquido que contiene surfactante y que tapiza el alvéolo, lo que reduce la tensión superficial del líquido alveolar. 2. El epitelio alveolar, que está formado por células epiteliales delgadas. 3. Una membrana basal epitelial. 4. Un espacio intersticial delgado entre el epitelio alveolar y la membrana capilar. 5. Una membrana basal capilar que en muchos casos se fusiona con la membrana basal del epitelio alveolar. 6. La membrana del endotelio capilar.
FACTORES QUE INFLUYEN EN LA VELOCIDAD DE DIFUSIÓN GASEOSA A TRAVÉS DE LA MEMBRANA RESPIRATORIA
Los factores que determinan la rapidez con la que un gas atraviesa la membrana son:
1) el grosor de la membrana.
2) el área superficial de la membrana.
3) el coeficiente de difusión del gas en la sustancia de la membrana.
y 4) la diferencia de presión parcial del gas entre los dos lados de la membrana.
El área superficial de la membrana respiratoria se puede reducir mucho en muchas situaciones. Por ejemplo, la resección de todo un pulmón reduce el área superficial total a la mitad de lo normal.
El coeficiente de difusión para la transferencia de cada uno de los gases a través de la membrana respiratoria depende de la solubilidad del gas en la membrana e inversamente de la raíz cuadrada del peso molecular del gas.
La diferencia de presión a través de la membrana respiratoria es la diferencia entre la presión parcial del gas en los alvéolos y la presión parcial del gas en la sangre capilar pulmonar.
Cuando la presión parcial de un gas en los alvéolos es mayor que la presión del gas en la sangre, como ocurre en el caso del O2, se produce difusión neta desde los alvéolos hacia la sangre; cuando la presión del gas en la sangre es mayor que la presión parcial en los alvéolos, como ocurre en el caso del CO2, se produce difusión neta desde la sangre hacia los alvéolos.
CAPACIDAD DE DIFUSIÓN DE LA MEMBRANA RESPIRATORIA
La capacidad de la membrana respiratoria de intercambiar un gas entre los alvéolos y la sangre pulmonar se expresa en términos cuantitativos por la capacidad de difusión de la membrana respiratoria, que se define como el volumen de un gas que difunde a través de la membrana en cada minuto para una diferencia de presión parcial de 1 mmHg. Todos los factores que se han analizado antes y que influyen en la difusión a través de la membrana respiratoria pueden influir sobre esta capacidad de difusión.
El pulmón tiene dos circulaciones: una circulación de bajo flujo y alta presión y una circulación de alto flujo y baja presión.La circulación de bajo flujo y alta presión aporta la sangre arterial sistémica a la tráquea, el árbol bronquial incluidos los bronquíolos terminales, los tejidos de sostén del pulmón y las capas exteriores (adventicias) de las arterias y venas pulmonares.
Las arterias bronquiales, que son ramas de la aorta torácica, irrigan la mayoría de esta sangre arterial sistémica a una presión solo ligeramente inferior a la presión aórtica.
La circulación de alto flujo y baja presiónque suministra la sangre venosa de todas las partes del organismo a los capilares alveolares en los que se añade el oxígeno (O2) y se extrae el dióxido de carbono (CO2).
La arteria pulmonar,que recibe sangre del ventrículo derecho, y sus ramas arteriales transportan sangre a los capilares alveolares para el intercambio gaseoso y a las venas pulmonares y después devuelven la sangre a la aurícula izquierda para su bombeo por el ventrículo izquierdo a través de la circulación sistémica.
VASOS PULMONARES
La arteria pulmonar se extiende solo 5 cm más allá de la punta del ventrículo derecho y después se divide en las ramas principales derecha e izquierda, que vascularizan los dos pulmones correspondientes.
La arteria pulmonar tiene un grosor de pared un tercio del de la aorta. Las ramas de las arterias pulmonares son cortas, y todas las arterias pulmonares, incluso las arterias más pequeñas y las arteriolas, tienen diámetros mayores que sus correspondientes arterias sistémicas. Este aspecto, combinado con el hecho de que los vasos son delgados y distensibles, da al árbol arterial pulmonar una gran distensibilidad, que es en promedio de casi 7 ml/mmHg, que es similar a la de todo el árbol arterial sistémico.
VASOS BRONQUIALES
La sangre también fluye hacia los pulmones a través de arterias bronquiales pequeñas que se originan en la circulación sistémica y transportan el 1-2% del gasto cardíaco total. Esta sangre arterial bronquial es sangre oxigenada, al contrario de la sangre parcialmente desoxigenada de las arterias pulmonares. Vascularizan los tejidos de soporte de los pulmones, como el tejido conjuntivo, los tabiques y los bronquios grandes y pequeños.
LINFÁTICOS
Hay vasos linfáticos en todos los tejidos de soporte del pulmón, comenzando en los espacios tisulares conjuntivos que rodean a los bronquíolos terminales, y siguiendo hacia el hilio del pulmón, y desde aquí principalmente hacia el conducto linfático torácico derecho.
CURSO DE PRESIÓN DEL VENTRÍCULO DERECHO
Estas curvas se comparan con la curva de presión aórtica, que es mucho más elevada, y que se muestra en la porción superior de la figura. La presión sistólica del ventrículo derecho del ser humano normal es en promedio de aproximadamente 25 mmHg, y la presión diastólica es en promedio de aproximadamente 0 a 1 mmHg, valores que son solo un quinto de los del ventrículo izquierdo.
PRESIONES EN LA ARTERIA PULMONAR
Durante la sístole la presión en la arteria pulmonar es esencialmente igual a la presión que hay en el ventrículo derecho, como también se muestra en la figura 39-1. Sin embargo, después del cierre de la válvula pulmonar al final de la sístole, la presión ventricular cae súbitamente, mientras que la presión arterial pulmonar disminuye más lentamente a medida que la sangre fluye a través de los capilares de los pulmones.
PRESIONES AURICULAR IZQUIERDA Y VENOSA PULMONAR
La presión media en la aurícula izquierda y en las venas pulmonares principales es en promedio de aproximadamente 2 mmHg en el ser humano en decúbito, y varía desde un valor tan bajo como 1 mmHg hasta uno tan elevado como 5 mmHg.
Habitualmente no es posible medir la presión auricular izquierda de un ser humano utilizando un dispositivo de medida directa porque es difícil introducir un catéter a través de las cavidades cardíacas hacia la aurícula izquierda. Sin embargo, con frecuencia se puede estimar la presión auricular izquierda con una exactitud moderada midiendo la denominada presión de enclavamiento pulmonar.
VOLUMEN SANGUÍNEO DE LOS PULMONES
El volumen de la sangre de los pulmones es de aproximadamente 450 ml, aproximadamente el 9% del volumen de sangre total de todo el aparato circulatorio. Aproximadamente 70 ml de este volumen de sangre pulmonar están en los capilares pulmonares, y el resto se divide aproximadamente por igual entre las arterias y las venas pulmonares.
LOS PULMONES SIRVEN COMO RESERVORIO DE SANGRE
En varias situaciones fisiológicas y patológicas la cantidad de sangre de los pulmones puede variar desde tan poco como la mitad del valor normal hasta el doble de lo normal. Por ejemplo, cuando una persona sopla aire con tanta intensidad que se genera una presión elevada en los pulmones (como cuando se toca una trompeta), se pueden expulsar hasta 250 ml de sangre desde el aparato circulatorio pulmonar hacia la circulación sistémica.
LA PATOLOGÍA CARDÍACA PUEDE DESPLAZAR SANGRE DESDE LA CIRCULACIÓN SISTÉMICA A LA CIRCULACIÓN PULMONAR
La insuficiencia del lado izquierdo del corazón o el aumento de la resistencia al flujo sanguíneo a través de la válvula mitral como consecuencia de una estenosis mitral o una insuficiencia mitral hace que la sangre quede estancada en la circulación pulmonar, aumentando a veces el volumen de sangre pulmonar hasta un 100% y produciendo grandes aumentos de las presiones vasculares pulmonares.
Dado que el volumen de la circulación sistémica es aproximadamente nueve veces el de la circulación pulmonar, el desplazamiento de sangre desde un sistema hacia el otro afecta mucho al sistema pulmonar, pero habitualmente tiene solo efectos circulatorios sistémicos leves.
LA DISMINUCIÓN DEL OXÍGENO ALVEOLAR REDUCE EL FLUJO SANGUÍNEO ALVEOLAR LOCAL Y REGULA LA DISTRIBUCIÓN DEL FLUJO SANGUÍNEO PULMONAR
Cuando la concentración de O2 en el aire de los alvéolos disminuye por debajo de lo normal (especialmente cuando disminuye por debajo del 70% de lo normal [es decir, por debajo de 73 mmHg de Po2]) los vasos sanguíneos adyacentes se constriñen, con un aumento de la resistencia vascular de más de cinco veces a concentraciones de O2 muy bajas.
Este efecto es opuesto al efecto que se observa en los vasos sistémicos, que se dilatan en lugar de constreñirse en respuesta a concentraciones bajas de O2.
Aunque los mecanismos que promueven la vasoconstricción pulmonar durante la hipoxia no se conocen en profundidad, la baja concentración de O2 puede estimular la liberación de sustancias vasoconstrictoras o reducir la liberación de un vasodilatador, como el óxido nítrico, del tejido pulmonar.
EFECTO DE LOS GRADIENTES DE PRESIÓN HIDROSTÁTICA DE LOS PULMONES SOBRE EL FLUJO SANGUÍNEO PULMONAR REGIONAL
La presión arterial en el pie de una persona que está de pie puede ser hasta 90 mmHg mayor que la presión a nivel del corazón. Esta diferencia está producida por la presión hidrostática, es decir, el peso de la propia sangre en los vasos sanguíneos. El mismo efecto, aunque en un grado menor, ocurre en los pulmones.
En el adulto en posición erguida el punto más bajo de los pulmones está normalmente unos 30 cm por debajo del punto más alto, lo que representa una diferencia de presión de 23 mmHg, de los cuales aproximadamente 15 mmHg están por encima del corazón y 8 por debajo. Es decir, la presión arterial pulmonar en la porción más elevada del pulmón de una persona que está de pie es aproximadamente 15 mmHg menor que la presión arterial pulmonar a nivel del corazón, y la presión en la porción más inferior de los pulmones es aproximadamente 8 mmHg mayor.
ZONAS 1, 2 Y 3 DEL FLUJO SANGUÍNEO PULMONAR
Los capilares de las paredes alveolares están distendidos por la presión de la sangre que hay en su interior, pero simultáneamente están comprimidos por la presión del aire alveolar que está en su exterior. Por tanto, siempre que la presión del aire alveolar pulmonar sea mayor que la presión de la sangre capilar, los capilares se cierran y no hay flujo sanguíneo. En diferentes situaciones normales y patológicas se puede encontrar una cualquiera de tres posibles zonas (patrones) del flujo sanguíneo pulmonar, como se señala a continuación:
Zona 1: ausencia de flujo durante todas las porciones del ciclo cardíaco porque la presión capilar alveolar local en esa zona del pulmón nunca aumenta por encima de la presión del aire alveolar en ninguna fase del ciclo cardíaco.
Zona 2: flujo sanguíneo intermitente,solo durante los picos de presión arterial pulmonar, porque la presión sistólica en ese momento es mayor que la presión del aire alveolar, pero la presión diastólica es menor que la presión del aire alveolar.
Zona 3: flujo de sangre continuo,porque la presión capilar alveolar es mayor que la presión del aire alveolar durante todo el ciclo cardíaco.
EL FLUJO SANGUÍNEO DE ZONA 1 SOLO SE PRODUCE EN SITUACIONES ANORMALES
El flujo sanguíneo de zona 1, que indica la ausencia de flujo durante todo el ciclo cardíaco, se produce cuando la presión arterial sistólica pulmonar es demasiado baja o cuando la presión alveolar es demasiado elevada para permitir que haya flujo. Por ejemplo, si una persona en posición erguida está respirando contra una presión aérea positiva de modo que la presión del aire intraalveolar es al menos 10 mmHg mayor de lo normal, pero la presión sanguínea sistólica pulmonar es normal, se puede esperar que se produzca flujo sanguíneo de zona 1 (ausencia de flujo sanguíneo) en los vértices pulmonares.
EL AUMENTO DEL GASTO CARDÍACO DURANTE EL EJERCICIO INTENSO ASUMIDO NORMALMENTE POR LA CIRCULACIÓN PULMONAR SIN GRANDES AUMENTOS EN LA PRESIÓN ARTERIAL PULMONAR
Durante el ejercicio intenso el flujo sanguíneo a través de los pulmones puede aumentar entre cuatro y siete veces. Este flujo adicional se acomoda en los pulmones de tres formas:
1) aumentando el número de capilares abiertos, a veces hasta tres veces.
2) distendiendo todos los capilares y aumentando la velocidad del flujo a través de cada capilar a más del doble.
y 3) aumentando la presión arterial pulmonar.
FUNCIÓN DE LA CIRCULACIÓN PULMONAR CUANDO LA PRESIÓN AURICULAR IZQUIERDA SE ELEVA COMO CONSECUENCIA DE UNA INSUFICIENCIA CARDÍACA IZQUIERDA
La presión auricular izquierda de una persona sana casi nunca se eleva por encima de +6 mmHg, incluso durante el ejercicio más intenso. Estas pequeñas modificaciones de la presión auricular izquierda prácticamente no tienen ningún efecto sobre la función de la circulación pulmonar porque simplemente expanden las vénulas pulmonares y abren más capilares, de modo que la sangre sigue fluyendo con una facilidad casi igual desde las arterias pulmonares.
Sin embargo, cuando se produce insuficiencia del lado izquierdo del corazón la sangre comienza a acumularse en la aurícula izquierda. Como consecuencia, la presión auricular izquierda puede aumentar de manera ocasional desde su valor normal de 1 a 5 mmHg hasta 40 a 50 mmHg. La elevación inicial de la presión auricular, de hasta aproximadamente 7 mmHg, tiene poco efecto sobre la función de la circulación pulmonar.
Sin embargo, cuando la presión auricular izquierda aumenta a más de 7 u 8 mmHg, aumentos adicionales de la presión auricular izquierda producen aumentos casi igual de grandes de la presión arterial pulmonar, generando de esta manera un aumento asociado de la carga del corazón derecho.
DINÁMICA CAPILAR PULMONAR
El intercambio de gases entre el aire alveolar y la sangre capilar pulmonar se analiza en el capítulo 40. Sin embargo, es importante señalar aquí que las paredes alveolares están tapizadas por tantos capilares que en la mayor parte de los sitios los capilares casi se tocan entre sí, adosados unos a otros. Por tanto, con frecuencia se dice que la sangre capilar fluye en las paredes alveolares como una «lámina de flujo», y no como capilares individuales.
INTERCAMBIO CAPILAR DE LÍQUIDO EN LOS PULMONES Y DINÁMICA DEL LÍQUIDO INTERSTICIAL PULMONAR
La dinámica del intercambio de líquido a través de las membranas capilares pulmonares escualitativamente la misma que en los tejidos periféricos. Sin embargo, cuantitativamente hay diferencias importantes, como se señala a continuación:
1. La presión capilar pulmonar es baja, de aproximadamente 7 mmHg, en comparación con una presión capilar funcional mucho mayor en los tejidos periféricos, de aproximadamente 17 mmHg.
2. La presión del líquido intersticial del pulmón es ligeramente más negativa que en el tejido subcutáneo periférico. (Esta presión se ha medido de dos formas: con una micropipeta insertada en el intersticio pulmonar, que da un valor de aproximadamente –5 mmHg, y midiendo la presión de absorción de líquido desde los alvéolos, que da un valor de aproximadamente –8 mmHg.)
3. La presión coloidosmótica del líquido intersticial pulmonar es de aproximadamente 14 mmHg, en comparación con menos de la mitad de este valor en los tejidos periféricos.
4. Las paredes alveolares son muy delgadas, y el epitelio alveolar que recubre las superficies alveolares es tan débil que se puede romper si la presión positiva en los espacios intersticiales es mayor que la presión del aire alveolar (>0 mmHg), lo que permite el paso de líquido desde los espacios intersticiales hacia los alvéolos.
EDEMA PULMONAR
El edema pulmonar se produce de la misma forma en que se produce el edema en cualquier otra localización del cuerpo. Cualquier factor que aumente la filtración de líquido fuera de los capilares pulmonares o que impida la función linfática pulmonar y provoque un aumento de la presión del líquido intersticial pulmonar desde el intervalo negativo hasta el intervalo positivo dará lugar al llenado rápido de los espacios intersticiales pulmonares y de los alvéolos con grandes cantidades de líquido libre.
Las causas más frecuentes de edema pulmonar son:
Insuficiencia cardíaca izquierda o valvulopatía mitral, con los consiguientes grandes aumentos de lapresión venosa pulmonar y de la presión capilar pulmonar y el encharcamiento de los espaciosintersticiales y de los alvéolos.
La lesión de las membranas de los capilares sanguíneos pulmonares producida por infecciones comola neumonía o por la inhalación de sustancias tóxicas como el gas cloro o el gas dióxido de azufre. Cada uno de estos mecanismos da lugar a una fuga rápida tanto de proteínas plasmáticas como de líquido desde los capilares hacia los espacios intersticiales pulmonares y los alvéolos.
LIQUIDO EN LA CAVIDAD PLEURAL
Cuando los pulmones se expanden y se contraen durante la respiración normal se deslizan en el interior de la cavidad pleural. Para facilitar este movimiento hay una delgada capa de líquido mucoide entre las pleuras parietal y visceral.
La membrana pleural es una membrana serosa mesenquimatosa porosa a través de la cual trasudan continuamente pequeñas cantidades de líquido intersticial hacia el espacio pleural. Estos líquidos arrastran con ellos proteínas tisulares, lo que da al líquido pleural una característica mucoide, que es lo que permite el deslizamiento muy fácil de los pulmones en movimiento.
PRESIÓN NEGATIVA EN EL LÍQUIDO PLEURAL
Siempre es necesaria una fuerza negativa en el exterior de los pulmones para mantener expandidos los pulmones. Esta fuerza es proporcionada por la presión negativa del espacio pleural normal.
La causa básica de esta presión negativa es el bombeo de líquidos desde el espacio pleural por los linfáticos (que también es la base de la presión negativa que se encuentra en la mayor parte de los espacios tisulares del cuerpo).
Como la tendencia al colapso normal de los pulmones es de aproximadamente –4 mmHg, la presión del líquido pleural siempre debe ser al menos tan negativa como –4 mmHg para mantener expandidos los pulmones.
DERRAME PLEURAL: ACUMULACIÓN DE GRANDES CANTIDADES DE LÍQUIDO LIBRE EN EL ESPACIO PLEURAL
El derrame es análogo al líquido de edema en los tejidos y se puede denominar «edema de la cavidad pleural». Las causas del derrame son las mismas que las causas del edema en otros tejidos, entre ellas:
1) bloqueo del drenaje linfático desde la cavidad pleural.
2) insuficiencia cardíaca, que da lugar a unas presiones capilares periférica y pulmonar excesivamente altas, que dan lugar a una trasudación excesiva de líquido hacia la cavidad pleural.
3) marcada reducción de la presión osmótica coloidal del plasma, que permite una trasudación excesiva de líquidos.
y 4) infección o cualquier otra causa de inflamación de las superficies de la cavidad pleural, que aumenta la permeabilidad de las membranas capilares y permite la salida rápida tanto de proteínas plasmáticas como de líquido hacia la cavidad
Las funciones principales de la respiración son proporcionar oxígeno a los tejidos y retirar el dióxido de carbono, realizado mediante 4 componentes principales:
1.- Ventilación pulmonar: flujo de entrada y salida de aire entre la atmósfera y los alvéolos pulmonares.
2.- Difusión de oxígeno (O2) y de dióxido de carbono (Co2) entre los alvéolos y la sangre.
3.- Transporte de oxígeno y de dióxido de carbono en la sangre y los líquidos corporales hacia las células de los tejidos corporales y desde las mismas.
4.- Y regulación de la ventilación y otras facetas de la respiración.
MÚSCULOS QUE CAUSAN LA EXPANSIÓN Y CONTRACCIÓN PULMONAR
Los pulmones se pueden expandir y contraer de dos maneras:
1.- Mediante el movimiento hacia arriba y hacia abajo del diafragma para alargar o acortar la cavidad torácica.
2.- Y mediante la elevación y el descenso de las costillas para aumentar o reducir el diámetro anteroposterior de la cavidad torácica.
La respiración tranquila normal se consigue por el primer mecanismo, es decir, por el movimiento del diafragma. Durante la inspiración la contracción del diafragma tira de las superficies inferiores de los pulmones, después durante la espiración, el diafragma simplemente se relaja, y el retroceso elástico de los pulmones, cavidad torácica y estructuras abdominales comprimen los pulmones de manera que se expulsa el aire hacia el exterior.
Sin embargo durante la respiración forzada las fuerzas elásticas no son lo suficientemente potentes como para expulsar el aire tan rápido como se inspira, de manera que se necesita una fuerza adicional que será aportada por los músculos abdominales que empujaran el contenido abdominal hacia arriba contra las superficies inferiores de los pulmones, comprimiéndolos de esta forma.
El segundo mecanismo para producir la respiración se basa principalmente en elevar la caja torácica. Esto se logra a través de la acción de una serie de músculos que permitirán la inspiración y espiración, es decir, se dividirán en músculos inspiratorios y espiratorios, los cuales se mencionan a continuación:
Músculos inspiratorios (elevadores de la caja torácica):
1.- Músculos intercostales externos.
2.- Músculos esternocleidomastoideos: elevan el esternón.
4.- Músculos escálenos: elevan las 2 primeras costillas.
Músculos espiratorios (desciendes la caja torácica):
1.- Músculos rectos del abdomen: empujan hacia abajo las costillas inferiores.
2.- Músculos intercostales internos.
PRESIÓN PLEURAL Y SUS CAMBIOS DURANTE LA INSPIRACIÓN
Es la presión de líquido que está en el delgado espacio comprendido entre la pleura pulmonar y la pleura de la cavidad torácica.
Sin embargo, debido a que es una aspiración ligera, es normalmente una presión negativa.
La presión pleural normal al comienzo de la inspiración es de -5cmH2O, lo suficiente para mantener expandido al pulmón hasta su posición de reposo, y durante la inspiración normal, la expansión de la caja torácica tira hacia afuera a los pulmones con más fuerza y genera una presión pleura aun mas negativa, de modo que alcanza una presión de -7.5cmH2O
PRESIÓN ALVEOLAR: PRESIÓN DEL AIRE EN EL INTERIOR DE LOS ALVÉOLOS PULMONARES
Cuando la glotis está abierta y no hay flujo de aire hacia el interior ni el exterior de los pulmones, las presiones en todas las partes del árbol respiratorio, hasta los alvéolos, son iguales a la presión atmosférica, que se considera que es la presión de referencia cero en las vías aéreas (es decir, presión de 0 cmH2O).
Existe una ligera presión negativa que es suficiente para arrastrar 0,5 l de aire hacia los pulmones en los 2 s necesarios para una inspiración tranquila normal. Durante la espiración, la presión alveolar aumenta hasta aproximadamente +1 cmH2O, lo que fuerza la salida del 0,5 l de aire inspirado desde los pulmones durante los 2 a 3 s de la espiración.
PRESIÓN TRANSPULMONAR: DEIFERENCIAS ENTRE LAS PRESIONES ALVEOLAR Y PLEURAL
la presión transpulmonar es la diferencia entre la presión que hay en el interior de los alvéolos y la que hay en las superficies externas de los pulmones (presión pleural), y es una medida de las fuerzas elásticas de los pulmones que tienden a colapsarlos en todos los momentos de la respiración, denominadas presión de retroceso.
DISTENSIBILIDAD DE LOS PULMONES
El volumen que se expanden los pulmones por cada aumento unitario de presión transpulmonar (si se da tiempo suficiente para alcanzar el equilibrio) se denomina distensibilidad pulmonar.
La distensibilidad pulmonar total de los dos pulmones en conjunto en el ser humano adulto normal es en promedio de aproximadamente 200 ml de aire por cada cmH2O de presión transpulmonar.
DIAGRAMA DE DISTENSIBILIDAD DE LOS PULMONES
es un diagrama que relaciona los cambios del volumen pulmonar con los cambios de la presión pleural, lo que, a su vez, modifica la presión transpulmonar.
En cada una de las curvas se registra modificando la presión pleural en escalones pequeños y permitiendo que el volumen pulmonar llegue a un nivel estable entre escalones sucesivos. Las dos curvas se denominan, respectivamente, la curva de distensibilidad inspiratoria y la curva de distensibilidad espiratoria y todo el diagrama se denomina diagrama de distensibilidad de los pulmones.
Las características del diagrama de distensibilidad están determinadas por las fuerzas elásticas de los pulmones. Estas se pueden dividir en dos partes:
1) fuerzas elásticas del tejido pulmonaren sí mismo,
y 2) fuerzas elásticas producidas por la tensión superficial del líquido que tapiza las paredes internas de los alvéolos y de otros espacios aéreos pulmonares.
PRINCIPIO DE LA TENSIÓN SUPERFICIAL
Cuando el agua forma una superficie con el aire, las moléculas de agua de la superficie del agua tienen una atracción especialmente intensa entre sí. En consecuencia, la superficie del agua siempre está intentando contraerse. Esto es lo que mantiene unidas entre sí las gotas de lluvia: una membrana muy contráctil de moléculas de agua que rodea toda la superficie de la gota de agua.
EL SUFACTANTE Y SU EFECTO SOBRE LA TENSIÓN SUPERFICIAL
El surfactante es un agente activo de superficie en agua, lo que significa que reduce mucho la tensión superficial del agua. Es secretado por células epiteliales especiales secretoras de surfactante denominadas células epiteliales alveolares de tipo II, que constituyen aproximadamente el 10% del área superficial de los alvéolos.
El surfactante es una mezcla compleja de varios fosfolípidos, proteínas e iones. Los componentes más importantes son el fosfolípido dipalmitoilfosfatidilcolina, las apoproteínas del surfactante e iones calcio.
En términos cuantitativos la tensión superficial de diferentes líquidos en agua es aproximadamente la siguiente: agua pura, 72 dinas/cm; los líquidos normales que tapizan los alvéolos pero sin surfactante, 50 dinas/cm; los líquidos normales que tapizan los alvéolos con cantidades normales de surfactante incluidas, entre 5 y 30 dinas/cm.
DISTENSIBILIDAD DEL TORÁX Y DE LOS PULMONES EN CONJUNTO
La distensibilidad de todo el sistema pulmonar (los pulmones y la caja torácica en conjunto) se mide cuando se expanden los pulmones de una persona relajada o paralizada totalmente.
Para medir la distensibilidad se introduce aire en los pulmones poco a poco mientras se registran las presiones y volúmenes pulmonares. Para insuflar este sistema pulmonar total es necesario casi el doble de presión que para insuflar los mismos pulmones después de extraerlos de la caja torácica.
Por tanto, la distensibilidad del sistema pulmón-tórax combinado es casi exactamente la mitad que la de los pulmones solos, 110 ml de volumen por cada cmH2O de presión para el sistema combinado, en comparación con200 ml/cmH2O para los pulmones de manera aislada.
REGISTRO DE LAS VARIACIONES DEL VOLUMEN PULMONAR: ESPIROMETRÍA
La ventilación pulmonar puede estudiarse registrando el movimiento del volumen del aire que entra y sale de los pulmones, un método que se denomina espirometría.
Un espirómetro básico típico, está formado por un tambor invertido sobre una cámara de agua, con el tambor equilibrado por un peso. En el tambor hay un gas respiratorio, habitualmente aire u oxígeno; un tubo conecta la boca con la cámara de gas. Cuando se respira hacia el interior y el exterior de la cámara, el tambor se eleva y desciende, y se hace un registro adecuado en una hoja de papel en movimiento.
Para facilitar la descripción de los acontecimientos de la ventilación pulmonar, el aire de los pulmones se ha subdividido en este diagrama en cuatro volúmenes y cuatro capacidades, que son el promedio de un hombre adulto joven.
VALORES MEDIOS DE LOS VOLÚMENES Y CAPACIDADES PULMONARES PARA UN HOMBRE ADULTO JOVEN Y SANO
VOLÚMENES PULMONARES
1. El volumen corrientees el volumen de aire que se inspira o se espira en cada respiración normal; es igual a aproximadamente 500 ml en el hombre adulto medio. 2. El volumen de reserva inspiratoriaes el volumen adicional de aire que se puede inspirar desde un volumen corriente normal y por encima del mismo cuando la persona inspira con una fuerza plena; habitualmente es igual a aproximadamente 3.000 ml. 3. El volumen de reserva espiratoria es el volumen adicional máximo de aire que se puede espirar mediante una espiración forzada después del final de una espiración a volumen corriente normal; normalmente, este volumen es igual a aproximadamente 1.100 ml. 4. El volumen residuales el volumen de aire que queda en los pulmones después de la espiración más forzada; este volumen es en promedio de aproximadamente 1.200 ml.
CAPACIDADES PULMONARES
1. La capacidad inspiratoriaes igual al volumen corriente más el volumen de reserva inspiratoria. Esta capacidad es la cantidad de aire (aproximadamente 3.500 ml) que una persona puede inspirar, comenzando en el nivel espiratorio normal y distendiendo los pulmones hasta la máxima cantidad. 2. La capacidad residual funcionales igual al volumen de reserva espiratoria más el volumen residual. Esta capacidad es la cantidad de aire que queda en los pulmones al final de una espiración normal (aproximadamente 2.300 ml).
3. La capacidad vital es igual al volumen de reserva inspiratoriamás el volumen corriente más elvolumen de reserva espiratoria. Esta capacidad es la cantidad máxima de aire que puede expulsar una persona desde los pulmones después de llenar antes los pulmones hasta su máxima dimensión y después espirando la máxima cantidad (aproximadamente 4.600 ml).
4. La capacidad pulmonar totales el volumen máximo al que se pueden expandir los pulmones con el máximo esfuerzo posible (aproximadamente 5.800 ml); es igual a la capacidad vital más el volumen residual.
VENTILACIÓN ALVEOLAR
la función de la ventilación pulmonar es renovar continuamente el aire de las zonas de intercambio gaseoso de los pulmones, en las que el aire está próximo a la sangre pulmonar. Estas zonas incluyen los alvéolos, los sacos alveolares, los conductos alveolares y los bronquíolos respiratorios. La velocidad a la que llega a estas zonas el aire nuevo se denomina ventilación alveolar.
ESPACIO MUERTO Y SU EFECTO SOBRE LA VENTILACION ALVEOLAR
Parte del aire que respira una persona nunca llega a las zonas de intercambio gaseoso, sino que simplemente llena las vías aéreas en las que no se produce intercambio gaseoso, como la nariz, la faringe y la tráquea. Este aire se denomina aire del espacio muerto, porque no es útil para el intercambio gaseoso.
Durante la espiración se expulsa primero el aire del espacio muerto, antes de que el aire procedente de los alvéolos llegue a la atmósfera. Por tanto, el espacio muerto es muy desventajoso para retirar los gases espiratorios de los pulmones.
TRÁQUEA, BRONQUIOS Y BRONQUIOLOS
no de los desafíos más importantes en todas las vías aéreas es mantenerlas abiertas y permitir el paso sin interrupciones de aire hacia los alvéolos y desde los mismos. Para evitar que la tráquea se colapse, múltiples anillos cartilaginosos se extienden aproximadamente 5/6 del contorno de la tráquea.
En las paredes de los bronquios, placas curvas de cartílago menos extensas también mantienen una rigidez razonable, aunque permiten un movimiento suficiente para que los pulmones se expandan y se contraigan.
Estas placas se hacen cada vez menos extensas en las últimas generaciones de bronquios y han desaparecido en los bronquíolos, que habitualmente tienen diámetros inferiores a 1,5 mm. No se impide el colapso de los bronquíolos por la rigidez de sus paredes. Por el contrario, se mantienen expandidos principalmente por las mismas presiones transpulmonares que expanden los alvéolos. Es decir, cuando los alvéolos se dilatan, los bronquíolos también se dilatan, aunque no tanto.
PARED MUSCULAR DE LOS BRONQUIOS Y BRONQUÍOLOS Y SU CONTROL
n todas las zonas de la tráquea y de los bronquios que no están ocupadas por placas cartilaginosas las paredes están formadas principalmente por músculo liso. Además, las paredes de los bronquíolos están formadas casi totalmente por músculo liso, con la excepción del bronquíolo más terminal, denominado bronquíolo respiratorio, que está formado principalmente por epitelio pulmonar y su tejido fibroso subyacente más algunas fibras musculares lisas.
RESISTENCIA AL FLUJO AÉREO EN EL ÁRBOL BRONQUIAL
En condiciones respiratorias normales el aire fluye a través de las vías aéreas con tanta facilidad que es suficiente un gradiente de presión menor de 1 cmH2O desde los alvéolos a la atmósfera para generar un flujo aéreo suficiente para una respiración tranquila.
La máxima resistencia al flujo aéreo no se produce en las pequeñas vías aéreas de los bronquíolos terminales, sino en algunos de los bronquíolos y bronquios de mayor tamaño cerca de la tráquea. La razón de esta elevada resistencia es que hay relativamente pocos de estos bronquios de mayor tamaño en comparación con los aproximadamente 65.000 bronquíolos terminales en paralelo, a través de los cuales solo debe pasar una cantidad muy pequeña de aire.
CONTROL NERVIOSO Y LOCAL DE LA MUSCULATURA BROQUIOLAR: DILATACIÓN SIMPÁTICA DE LOS BROQUÍOLOS
El control directo de los bronquíolos por las fibras nerviosas simpáticas es relativamente débil porque pocas fibras de este tipo penetran hasta las porciones centrales del pulmón. Sin embargo, el árbol bronquial está muy expuesto a la noradrenalina y adrenalina que se liberan hacia la sangre por la estimulación simpática de la médula de las glándulas suprarrenales.
Estas dos hormonas (especialmente la adrenalina, debido a su mayor estimulación de los receptores β-adrenérgicos) producen dilatación del árbol bronquial.
CONSTRICCIÓN PARASIMPÁTICA DE LOS BRONQUIOLOS
Algunas fibras nerviosas parasimpáticas procedentes de los nervios vagos penetran en el parénquima pulmonar. Estos nervios secretan acetilcolina y, cuando son activados, producen una constricción leve a moderada de los bronquíolos.
Cuando una enfermedad como el asma ya ha producido un cierto grado de constricción bronquiolar, la estimulación nerviosa parasimpática adicional con frecuencia empeora la enfermedad. Cuando se produce esta situación, la administración de fármacos que bloquean los efectos de la acetilcolina, como atropina, a veces puede relajar las vías aéreas lo suficiente para aliviar la obstrucción.
LOS FACTORES SECRETORES LOCALES PUEDEN PRODUCIR CONTRICCIÓN BRONQUÍOLAR
Algunas sustancias que se forman en los pulmones tienen con frecuencia bastante actividad en la producción de constricción bronquiolar.
Dos de las más importantes de estas sustancias son la histamina y la sustancia de reacción lenta de la anafilaxia. Estas dos sustancias se liberan a nivel pulmonar por los mastocitos durante las reacciones alérgicas, sobre todo las provocadas por pólenes del aire. Por tanto, juegan un papel fundamental en la obstrucción de la vía aérea observada en el asma alérgica, sobre todo la sustancia de reacción lenta de la anafilaxia.
MOCO QUE RECUBRE LAS VÍAS AÉREAS Y ACCIÓN DE LOS CILIOS EN LA LIMPIEZA DE LAS VÍAS AÉREAS
Todas las vías aéreas, desde la nariz a los bronquíolos terminales, están humedecidas por una capa de moco que recubre toda la superficie.
El moco es secretado en parte por las células caliciformes mucosas individuales del recubrimiento epitelial de las vías aéreas y en parte por pequeñas glándulas submucosas. Además de mantener humedecidas las superficies, el moco atrapa partículas pequeñas que están en el aire inspirado e impide que la mayoría de estas partículas llegue a los alvéolos.
Toda la superficie de las vías aéreas, tanto en la nariz como en las vías inferiores hasta los bronquíolos terminales, está tapizada por un epitelio ciliado que tiene aproximadamente 200 cilios por cada una de las células epiteliales. Estos cilios baten continuamente a una frecuencia de 10 a 20 veces por segundo y la dirección de su «golpe de fuerza» siempre se dirige hacia la faringe.
Es decir, los cilios de los pulmones baten hacia arriba, mientras que los de la nariz baten hacia abajo. Este batido continuo hace que la cubierta de moco fluya lentamente, a una velocidad de algunos milímetros por minuto, hacia la faringe. Después el moco y las partículas que están atrapadas en el mismo son deglutidos o se expulsan hacia el exterior con la tos.