
Organización de la médula espinal para las funciones motoras.
Las señales sensitivas penetran en ella por las raíces sensitivas, también conocidas como raíces posteriores o dorsales. Después de entrar, cada una viaja hacia dos destinos diferentes: una rama del nervio sensitivo termina casi de inmediato en la sustancia gris de la médula y suscita reflejos medulares segmentarios de ámbito local y otros efectos a este nivel, y otra rama transmite sus impulsos hacia niveles más altos del sistema nervioso, es decir, las zonas superiores de la propia médula, el tronco del encéfalo o incluso la corteza cerebral.
MOTONEURONAS ANTERIORES.
En ellas nacen las fibras nerviosas que salen de la médula a través de las raíces anteriores e inervan directamente las fibras de los músculos esqueléticos. Estas neuronas son de dos tipos, motoneuronas α y motoneuronas γ.
Motoneuronas α
Las motoneuronas α dan origen a unas fibras nerviosas motoras grandes de tipo Aα, con un promedio de 14 μm de diámetro; a lo largo de su trayecto se ramifican muchas veces después de entrar en el músculo e inervan las grandes fibras musculares esqueléticas. La estimulación de una sola fibra nerviosa α excita de tres a varios cientos de fibras musculares esqueléticas a cualquier nivel, que en conjunto reciben el nombre de unidad motora.
Motoneuronas γ
Además de las motoneuronas α, que activan la contracción de las fibras musculares esqueléticas, hay otras motoneuronas γ mucho más pequeñas que están situadas en las astas anteriores de la médula espinal, cuyo número es más o menos la mitad que las anteriores. Estas células transmiten impulsos a través de unas fibras nerviosas motoras γ de tipo A (Aγ) mucho más pequeñas, con un diámetro medio de 5 μm, que van dirigidas hacia unas fibras del músculo esquelético especiales pequeñas llamadas fibras intrafusales.
Interneuronas
Las interneuronas están presentes en todas las regiones de la sustancia gris medular, en las astas posteriores, las astas anteriores y las zonas intermedias que quedan entre ellas.
Su tamaño es pequeño y poseen una naturaleza muy excitable, pues con frecuencia muestran una actividad espontánea capaz de emitir hasta 1.500 disparos por segundo. Las conexiones entre las interneuronas y las motoneuronas anteriores son las responsables de la mayoría de las funciones integradoras.

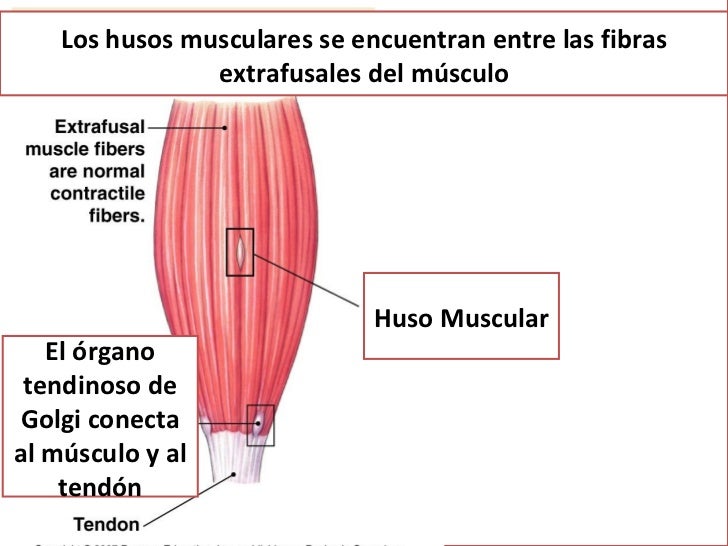
HUSO MUSCULAR
Cada elemento tiene una longitud de 3 a 10 mm. Se encuentra dispuesto alrededor de 3 a 12 fibras musculares intrafusales diminutas cuyos extremos acaban en punta y se fijan al glucocáliz de las grandes fibras extrafusales adyacentes correspondientes al músculo esquelético.
La porción receptora del huso muscular se localiza en su parte central.
Es fácil comprobar que el receptor del huso muscular puede excitarse por dos mecanismos:
1. El alargamiento del músculo en su conjunto estira la porción intermedia del huso y, por tanto, estimula al receptor.
2. Aunque la longitud de todo el músculo no cambie, la contracción de las porciones finales de las fibras intrafusales también estira la porción intermedia del huso y así activa el receptor.
TERMINACIÓN PRIMARIA.
En el centro de la zona receptora, una gran fibra nerviosa sensitiva rodea la porción central de cada fibra intrafusal, formando la denominada terminación aferente primaria o terminación anuloespiral. Esta fibra nerviosa es de tipo Ia, con un diámetro medio de 17 μm, y envía señales sensitivas hacia la médula espinal a una velocidad de 70 a 120 m/s, la mayor entre todos los tipos de fibras nerviosas en el cuerpo.
TERMINACIÓN SECUNDARIA.
La terminación receptora situada a un lado de la terminación primaria o a los dos normalmente está inervada por una fibra nerviosa sensitiva, pero a veces por dos más pequeñas (fibras de tipo II con un diámetro medio de 8 μm), tal como está representado en las figuras 55-3 y 55-4. Esta terminación sensitiva se llama terminación aferente secundaria; en ocasiones rodea a las fibras intrafusales de la misma forma que lo hace la fibra de tipo Ia, pero a menudo se extiende como las ramas de un arbusto.

Reflejo miotático muscular
La manifestación más sencilla del funcionamiento del huso es el reflejo miotático o de estiramiento muscular. Siempre que se estira bruscamente un músculo, la activación de los husos causa la contracción refleja de las fibras musculares esqueléticas grandes en el músculo estirado y también en los músculos sinérgicos más íntimamente ligados.

REFLEJO MIOTATICO DINÁMICO.
El reflejo miotático dinámico surge con potentes señales dinámicas transmitidas desde las terminaciones sensitivas primarias de los husos musculares, originada por su estiramiento o distensión rápida. Esto es, cuando un músculo se estira o se distiende bruscamente, se transmite un impulso potente hacia la médula espinal, lo que provoca instantáneamente una enérgica contracción refleja (o un descenso de la contracción) en el mismo músculo del que nació la señal. Por tanto, el reflejo sirve para oponerse a los cambios súbitos sufridos en la longitud muscular.
REFLEJO MIOTATICO ESTÁTICO.
Este reflejo deriva de las señales receptoras estáticas continuas transmitidas por las terminaciones primarias y secundarias. La importancia del reflejo miotático estático radica en que produce un grado de contracción muscular que puede mantenerse razonablemente constante, excepto cuando el sistema nervioso de la persona desee específicamente otra cosa
Intervención del huso muscular en la actividad motora voluntaria
Para comprender la importancia del sistema eferente γ es necesario saber que el 31% de todas las fibras nerviosas motoras dirigidas al músculo son fibras eferentes γ de tipo A pequeñas en vez de las fibras motoras α de tipo A grandes. Siempre que se transmiten señales desde la corteza motora o desde cualquier otra área del encéfalo hacia las motoneuronas α, las motoneuronas γ reciben un estímulo simultáneo en la mayoría de los casos, efecto denominado coactivación de las motoneuronas α y γ. Este efecto hace que se contraigan al mismo tiempo las fibras musculares esqueléticas extrafusales y las fibras intrafusales del huso muscular.

Áreas encefálicas que regulan el sistema motor γ.
El sistema eferente γ se activa de forma específica con las señales procedentes de la región facilitadora bulborreticular del tronco del encéfalo y, de un modo secundario, con los impulsos transmitidos hacia la zona bulborreticular desde: 1) el cerebelo; 2) los ganglios basales, y 3) la corteza cerebral
Aplicaciones clínicas del reflejo miotático.
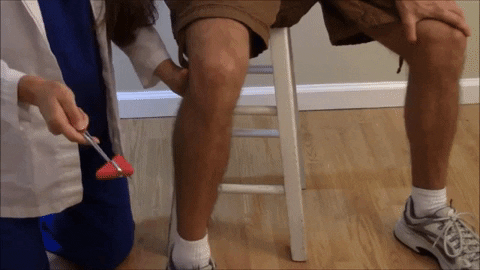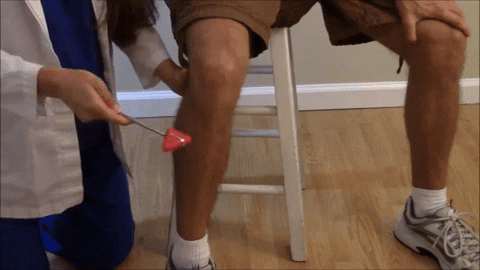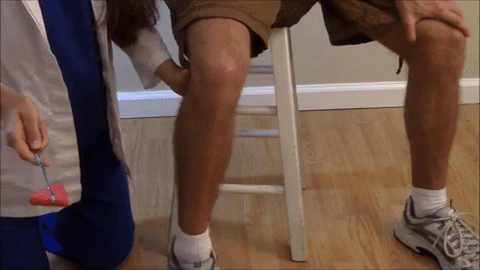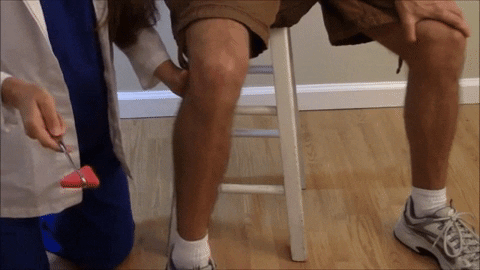
El reflejo rotuliano y otros reflejos de estiramiento muscular pueden usarse para valorar la sensibilidad de los reflejos miotáticos.
El reflejo rotuliano en concreto puede explorarse simplemente golpeando el tendón rotuliano con un martillo de reflejos; esta acción estira al instante el músculo cuádriceps y genera un reflejo miotático dinámico que hace que la pierna experimente una «sacudida» hacia delante.
Clono: oscilación de las sacudidas musculares
En ciertas condiciones, las sacudidas musculares pueden oscilar, fenómeno denominado clono. Cuando una persona de puntillas deja caer bruscamente su cuerpo hacia abajo y estira los músculos gastrocnemios, los impulsos del reflejo miotático se transmiten desde los husos musculares hacia la médula espinal. Estas señales excitan el músculo estirado de forma refleja, lo que vuelve a elevar el cuerpo. Al cabo de una fracción de segundo se extingue la contracción refleja del músculo y el cuerpo cae de nuevo, lo que estira los husos en una segunda oportunidad. Una vez más, un reflejo miotático dinámico levanta el cuerpo, pero en esta situación también se desvanece después de una fracción de segundo, y el cuerpo desciende de nuevo para comenzar el siguiente ciclo. De este modo, el reflejo miotático del músculo gastrocnemio sigue oscilando, a menudo durante largos períodos, que es un clono.
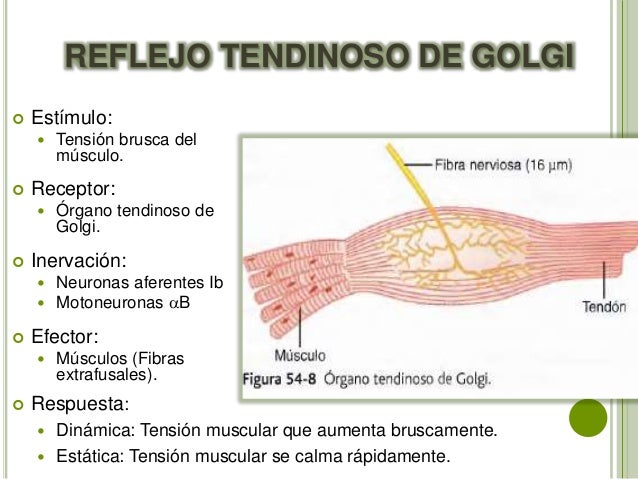
Reflejo tendinoso de Golgi
El órgano tendinoso de Golgi sirve para controlar la tensión muscular.Las señales procedentes del órgano tendinoso se transmiten a través de fibras nerviosas grandes de conducción rápida de tipo Ib, con un diámetro medio de 16 μm, tan solo un poco más pequeñas que las correspondientes a las terminaciones primarias del huso muscular. Tales fibras, igual que en el caso de estas últimas, envían impulsos hacia las zonas locales de la médula y, después de hacer sinapsis en el asta posterior, siguen a través de las vías de fibras largas, como los fascículos espinocerebelosos dirigidos hacia el cerebelo, y todavía a través de otros fascículos más hacia la corteza cerebral. Las señales medulares locales estimulan una sola interneurona inhibidora que actúa sobre la motoneurona anterior. Este circuito local inhibe directamente el músculo correspondiente sin influir sobre los músculos adyacentes. Otra probable función del reflejo tendinoso de Golgi consiste en igualar las fuerzas de contracción de las distintas fibras musculares. A saber, las fibras que ejerzan una tensión excesiva quedan inhibidas por su intervención, mientras que las que produzcan una tensión demasiado ligera reciben una mayor excitación debido a la ausencia de la inhibición refleja. Este fenómeno dispersa la carga muscular entre todas las fibras e impide la lesión de zonas aisladas de un músculo donde una pequeña cantidad de fibras pudiera verse sobrecargada
Reflejo flexor y reflejos de retirada.
En el animal espinal o descerebrado es fácil que prácticamente cualquier tipo de estímulo sensitivo cutáneo de los miembros haga que sus músculos flexores se contraigan, lo que permite retirar la extremidad del objeto estimulador. Es el denominado reflejo flexor. En su forma clásica, el reflejo flexor se suscita con mayor potencia mediante la estimulación de las terminaciones para el dolor, como sucede con un pinchazo, el calor o una herida, razón por la que también se le denomina reflejo nociceptivo, o simplemente reflejo al dolor. La activación de los receptores para el tacto también puede despertar un reflejo flexor más débil y menos prolongado. Si cualquier parte del cuerpo aparte de las extremidades recibe un estímulo doloroso, esa porción se alejará del estímulo en correspondencia, pero el reflejo puede no quedar limitado a los músculos flexores, aun cuando sea básicamente el mismo tipo de fenómeno. Por tanto, cualquiera de los múltiples patrones que adoptan en las diferentes regiones del organismo se llama reflejo de retirada.

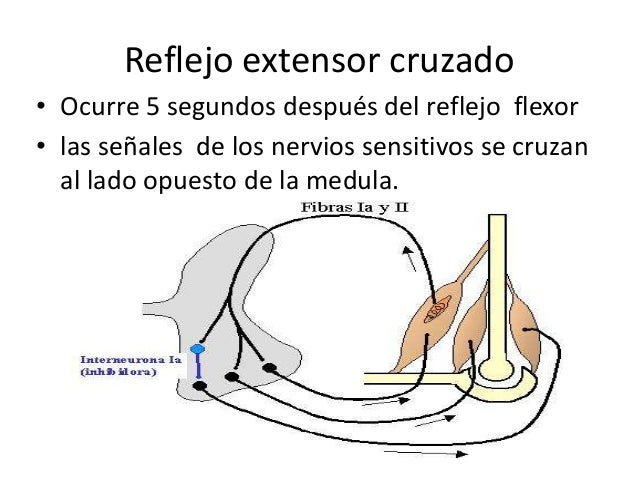
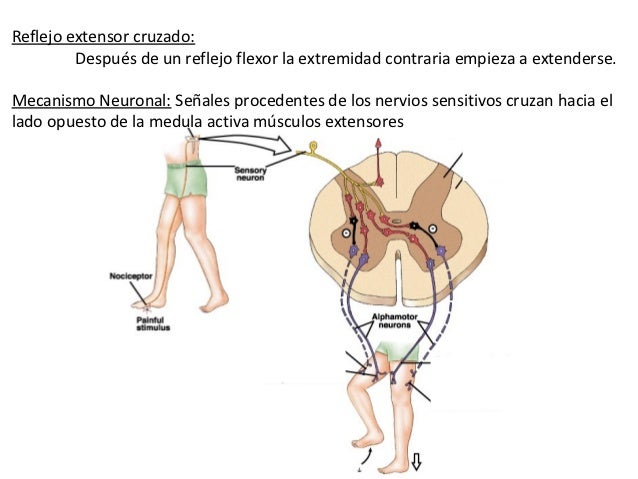
Reflejo extensor cruzado
Más o menos entre 0,2 y 0,5 s después de que cualquier estímulo suscite un reflejo flexor en una extremidad, la extremidad contraria comienza a extenderse. Este reflejo se denomina reflejo extensor cruzado. La extensión del miembro opuesto puede tirar de todo el cuerpo para alejarlo del objeto que origina el estímulo doloroso en el miembro apartado. Dado que este reflejo no suele comenzar hasta unos 200 a 500 ms después de haber comenzado el estímulo doloroso inicial, no hay duda de que en el circuito formado entre la neurona sensitiva aferente y las motoneuronas del lado contrario de la médula encargadas de la extensión cruzada participan muchas interneuronas. Una vez que ha desaparecido el estímulo doloroso, el reflejo extensor cruzado presenta un período de posdescarga aún más largo que en el caso del reflejo flexor. Una vez más, se cree que esta extensa posdescarga deriva de los circuitos reverberantes establecidos entre las interneuronas.
Inhibición e inervación recíprocas.
cuando un reflejo miotático activa un músculo, a menudo inhibe simultáneamente a sus antagonistas. Este es el fenómeno de la inhibición recíproca y el circuito neuronal que da lugar a una relación de este tipo se llama de inervación recíproca. En este mismo sentido, suelen existir relaciones recíprocas entre los músculos de los dos lados del cuerpo, tal como queda ejemplificado por los reflejos musculares flexores y extensores.
Reflejos posturales y locomotores.
Reacción de apoyo positiva.
La reacción de apoyo positiva implica un circuito de interneuronas complejo, semejante a los circuitos responsables de los reflejos flexor y extensor cruzado. El punto de presión sobre la almohadilla plantar determina la dirección con la que se extenderá el miembro; su aplicación sobre un lado causa la extensión en esa misma dirección, efecto denominado reacción del imán. Esta reacción sirve para impedir que el animal se caiga hacia ese lado.
Reflejos medulares de enderezamiento.
Cuando un animal espinal está tendido sobre su costado, realizará movimientos descoordinados para tratar de incorporarse. Es el denominado reflejo de enderezamiento medular. Dicho fenómeno pone de manifiesto que la integración de algunos reflejos relativamente complejos asociados a la postura tiene lugar en la médula espinal. En efecto, un animal con una médula torácica cortada y perfectamente cicatrizada entre los niveles de inervación para las patas anteriores y las posteriores puede enderezarse por sí solo desde su posición tumbada e incluso caminar con sus patas traseras además de las delanteras

Movimientos de la marcha y la deambulación.
Movimientos ritmicos de la marcha en un solo miembro.
Las señales sensitivas procedentes de las almohadillas plantares y de los sensores posturales que rodean a las articulaciones desempeñan un cometido relevante para controlar la presión aplicada sobre la pata y la frecuencia de los pasos cuando se la deja caminar a lo largo de una superficie. En realidad, el mecanismo medular para regular la marcha puede ser aún más complicado. Por ejemplo, si la parte superior de la pata tropieza con un obstáculo durante su propulsión hacia adelante, esta maniobra sufrirá una detención transitoria; a continuación, y según una rápida sucesión, la pata se alzará más alta y avanzará hacia adelante para superar el obstáculo. Este es el reflejo del tropezón. Por tanto, la médula representa un mecanismo controlador inteligente de la marcha
Marcha recíproca de las extremidades opuestas
Si la médula espinal lumbar no se secciona hasta el centro, cada vez que se den unos pasos en sentido hacia delante con una extremidad, la opuesta corrientemente se desplaza hacia atrás. Este efecto deriva de la inervación recíproca existente entre ambos miembros
Marcha en diagonal entre las cuatro extremidades: el reflejo de «marcar el paso»
Si se sostiene a un animal espinal bien restablecido (con una sección medular a nivel del cuello por encima de la zona destinada a la pata delantera en la médula) encima del suelo y se deja que sus patas se balanceen, el estiramiento de las extremidades a veces desencadena reflejos de la marcha en los que participan las cuatro patas. En general, los pasos siguen un patrón en diagonal entre las patas delanteras y las traseras. Esta respuesta diagonal constituye otra manifestación de la inervación recíproca, esta vez a lo largo de toda la longitud de la médula hacia arriba y hacia abajo entre las extremidades anteriores y las posteriores. Este patrón de marcha se denomina reflejo de marcar el paso
Reflejo de galope
Otro tipo de reflejo que a veces surge en un animal espinal es el reflejo de galope, en el que las extremidades anteriores se desplazan hacia atrás al unísono a la vez que las posteriores se mueven hacia delante. Este reflejo suele suceder cuando se aplican estímulos casi idénticos de estiramiento o de presión a las extremidades de ambos lados del cuerpo al mismo tiempo: su estimulación dispar promueve el reflejo de la marcha en diagonal. Esto encaja con los patrones normales de la marcha y el galope, porque al caminar, cada vez no se estimula nada más que una pata delantera y otra trasera, lo que pondría al animal en condiciones de seguir avanzando. En cambio, al golpear el suelo durante el galope, las dos extremidades anteriores y las dos posteriores se estimulan más o menos por igual, lo que le deja listo para continuar galopando y, por tanto, mantener este patrón de movimiento

Reflejo de rascado.
Un reflejo medular especialmente importante en algunos animales es el reflejo de rascado, que se pone en marcha cuando se percibe una sensación de prurito o de cosquilleo. Este reflejo abarca dos funciones: 1) una sensibilidad postural que permite a la garra o la zarpa encontrar el punto exacto de irritación sobre la superficie del cuerpo, y 2) un movimiento de vaivén para el rascado. La sensibilidad postural del reflejo de rascado es una función muy evolucionada. Si se mueve una pulga por una región tan anterior como el hombro de un animal espinal, la garra posterior aún es
capaz de encontrar este punto, pese a que para poder alcanzarlo han de contraerse 19 músculos a la vez en la extremidad según un patrón preciso
Espasmo muscular producido por una fractura ósea
En los músculos que rodean a un hueso fracturado aparece un tipo de espasmo importante desde el punto de vista clínico. El espasmo obedece a los impulsos dolorosos puestos en marcha desde los extremos del hueso roto, que hacen que los músculos en torno a esta zona experimenten una contracción tónica. El alivio del dolor obtenido mediante la inyección de un anestésico local en los bordes fragmentados del hueso atenúa el espasmo; la anestesia general profunda de todo el cuerpo, como por ejemplo el empleo de éter, también mitiga el espasmo.

Espasmo de la musculatura abdominal en personas con peritonitis.
Otro tipo de espasmo local ocasionado por los reflejos medulares es el espasmo abdominal resultante de la irritación experimentada por el peritoneo parietal en una peritonitis. Aquí de nuevo el alivio del dolor generado por la peritonitis permite la relajación del músculo espástico. El mismo tipo de espasmo sucede muchas veces en el curso de las intervenciones quirúrgicas; por ejemplo, en las operaciones abdominales, los impulsos dolorosos procedentes del peritoneo parietal suelen hacer que los músculos del abdomen se contraigan intensamente, lo que a veces expulsa los intestinos a través de la herida quirúrgica. Por esta razón, en la cirugía abdominal suele ser necesario recurrir a la anestesia profunda.
Calambres musculares.
Otro tipo de espasmo local es el típico calambre muscular. Cualquier factor local irritante o la perturbación metabólica de un músculo, como el frío intenso, la ausencia de flujo sanguíneo o el ejercicio excesivo, pueden despertar dolor u otras señales sensitivas que se transmitan desde el músculo hasta la médula espinal, y a su vez desencadenen una contracción refleja en el músculo como mecanismo de autorregulación. Se cree que la contracción estimula los mismos receptores sensitivos todavía más, lo que hace que la médula espinal acentúe la intensidad de la contracción. Por tanto, se produce una retroalimentación positiva, de modo que un pequeño nivel inicial de irritación origina una contracción cada vez mayor hasta que sobreviene un auténtico calambre muscular.
