El ojo II. Función receptora y nerviosa de la retina.

Retina
La retina es la capa mas interna del globo ocular. Su misión es transformar la luz que recibe, en un impulso nervioso que viaja hasta el cerebro a través del nervio óptico, y se convierte en las imágenes que percibimos. La luz llega a través de la córnea, cruzando la pupila y el cristalino hasta que llega a la retina. Es necesario que todas las estructuras estén sanas para una buena visión.

La retina está formada por entramado de células nerviosas que viajan hasta el cerebro por el nervio óptico. Esto hace que tengan una distribución anatómica especial en el fondo de ojo:

Papila óptica: es la cabeza del nervio óptico, donde se agrupan todos las prolongaciones de las neuronas para llevar el impuso hacia el cerebro.
Arteria y vena de la retina: Son las encargadas de nutrir las capas más internas de la retina.
Mácula: Es la zona de la retina encargada de la visión central, de la visión final de los detalles de las cosas (leer, enhebrar una aguja).
Fóvea: Es el centro de la visión, donde se enfocan los rayos de luz que llegan a la retina.
Retina periférica: Es la que nos permite la visión periférica. Si alguien está fuera de donde miramos, la retina periférica nos permite reconocerlo por su forma. Vítreo: Es el gel que ocupa la cavidad del globo ocular. Es transparente y está pegado a la retina. Con la edad va sufriendo proceso de envejecimiento y es el responsable de las “moscas volantes” que ven algunos pacientes.

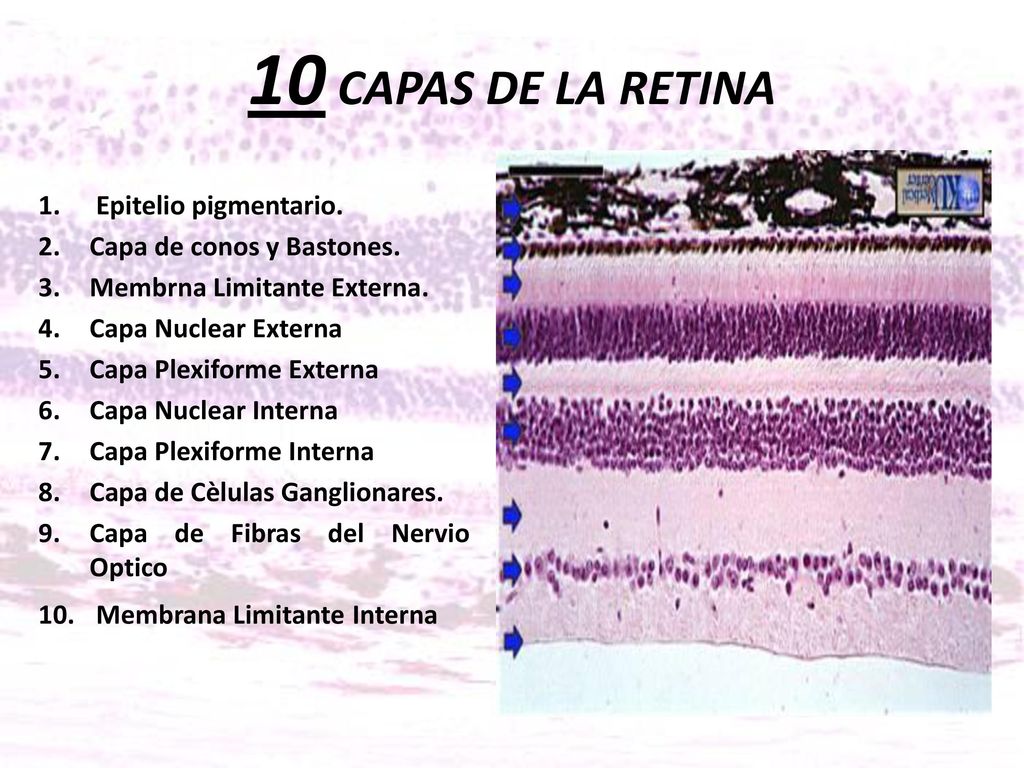
La retina está formada por diez capas bien diferenciadas.
Están íntimamente conectadas para ir transformando la señal de luz que reciben en el impulso nervioso. Las agruparemos en 3 secciones para hacerlo más comprensible.
Epitelio pigmentario y fotorreceptores: Los fotorreceptores son los encargadas de transformar la luz en seña eléctrica. Hay dos tipos:
Conos: funcionan con mucha luz, son las encargadas de la visión de los colores.
Bastones: responsables de la visión nocturna.
Células bipolares: Transmiten la señal hasta las células ganglionares.
Células ganglionares: Son la última capa. De ellas nacen las prolongaciones nerviosas que formaran el nervio óptico.
Sus 10 capas son :
1.Capa pigmentaria 2. Capa de conos y bastones 3. Membrana limitante externa 4. Capa nuclear externa 5. Capa plexiforme externa 6. Capa nuclear interna 7. Capa plexiforme interna 8. Capa ganglionar 9. Capa de fibras del nervio óptico 10. Membrana limitante interna

FOVEA CENTRAL
Especializada para la visión aguda y detallada, compuesta por conos en su integridad estos conos contribuyen a la detección de detalles en la imagen visual •Región central: quedan atrapadas las capas internas de la retina para atenuar la perdida de la agudeza

/231053764009/231053764009_gf30.png)
CONOS Y BASTONES

MELANINA
Impide la reflexión lumínica por toda la esfera del globo factor importante en la visión nítida Los rayos de luz se reflejarían en todas las direcciones dentro del Globo ocular y generarían una iluminación difusa de la retina en vez del contraste normal de los puntos claros y oscuros necesarios para la formación de una imagen precisa
20/100 20/200 normal:20/20



DESPRENDIMIENTO DE LA RETINA

Fotoquímica de la visión
Ciclo rodopsina-retinal y excitación de los bastones. La rodopsina se descompone por la energía lumínica. El fotopigmento rodopsina de los bastones se concentra en la porción del segmento externo quesobresale hacia la capa pigmentaria. Esta sustancia es una combinación de la proteína escotopsina y del pigmento carotenoide retinal o, más concretamente, 11-cis retinal. Cuando la rodopsina absorbe la energía lumínica, el retinal se transforma en su configuración todo-trans y comienzan a separarse los componentes retinal y escotopsina. Tras una serie de reacciones extraordinariamente rápidas, el componente retinal se transforma en lumirrodopsina, metarrodopsina I, metarrodopsina II y, por último, escotopsina; y se escinde el todo-trans retinal. Durante este proceso, se cree que la metarrodopsina II se ocupa de la transmisión de los impulsos sucesivos por la retina.
Regeneración de la rodopsina.
En la primera etapa de regeneración de la rodopsina, el todo-trans retinal se convierte en 11-cis retinal, que se combina en seguida con la escotopsina para dar la rodopsina. Existe una segunda vía de formación de la rodopsina basada en la conversión del todotrans retinal en todo-transretinol, unaformadevitaminaA.El retinol se convierte por vía enzimática en 11-cis retinol y luego en 11-cis retinal, que puede combinarse con la escotopsina para dar rodopsina. Si la retina contiene un exceso de retinal, se transforma en vitamina A, disminuyendo así la cantidad total de rodopsina de la retina. Las personas con carencia de vitamina A experimentan ceguera nocturna porque los bastones son los fotorreceptores que más se utilizan en condiciones de iluminación crepuscular y la formación de rodopsina disminuye de forma espectacular por la ausencia de vitamina A. Este trastorno se revierte en 1 h o menos mediante la inyección intravenosa de vitamina A.

Función de la vitamina A en la formación de la rodopsina.
existe una segunda vía química por la que el todo-trans-retinal puede convertirse en 11-cis-retinal. En este caso consiste en la transformación primero del todo -trans-retinal en todo-trans- retinol, que es una forma de la vitamina A. A continuación, el todo-trans-retinol pasa a 11-cis-retinol bajo la influencia de la enzima isomerasa. Finalmente, el 11-cis-retinol da lugar a 11-cis-retinal, que se combina con la escotopsina para generar nueva rodopsina. La vitamina A está presente en el citoplasma de los bastones y en la capa pigmentaria de la retina. Por tanto, normalmente siempre se encuentra disponible para producir nuevo retinal cuando haga falta. A la inversa, si hay un exceso de retinal en la retina, vuelve a transformarse en vitamina A, lo que reduce la cantidad de pigmento sensible a la luz que existe en ella. Más adelante veremos que esta interconversión entre el retinal y la vitamina A resulta especialmente importante para la adaptación prolongada de la retina a las diferentes intensidades de luz.


Ceguera nocturna.
La ceguera nocturna o hesperanopía aparece en una persona con una hipovitaminosis A grave. La razón para ello simplemente estriba en que, sin vitamina A, la cantidad de retinal y de rodopsina que puede formarse disminuye acusadamente. Esta situación se denomina ceguera nocturna porque la magnitud de luz existente por la noche es demasiado escasa como para permitir una visión suficiente en las personas aquejadas de una deficiencia de vitamina A. Para que surja una ceguera nocturna, las personas habitualmente deben seguir una alimentación desprovista de vitamina A durante meses, dado que en condiciones normales hay grandes cantidades almacenadas en el hígado, que pueden quedar a disposición de los ojos. Una vez que se contrae esta alteración, a veces puede desaparecer en menos de 1 h mediante la inyección intravenosa de vitamina A

Excitación del bastón cuando la luz activa la rodopsina
la excitación de un bastón provoca un aumento de la negatividad en el potencial de membrana en su interior, lo que supone un estado de hiperpolarización, que significa una mayor negatividad de la normal dentro de su membrana. Este fenómeno es exactamente lo contrario del descenso de la negatividad (el proceso de «despolarización») que sucede en casi todos los demás receptores sensitivos. Pero la activación de la rodopsina, ¿cómo suscita la hiperpolarización? La solución a esta pregunta dice que cuando se descompone la rodopsina, disminuye la conductancia de la membrana del bastón para los iones sodio en su segmento externo. Esto provoca una hiperpolarización de toda la membrana del bastón por el siguiente mecanismo.


Mecanismo por el que la descomposición de la rodopsina disminuye la conductancia al sodio de la membrana: la «cascada» de excitación.
En condiciones óptimas, un solo fotón de luz, la unidad cuántica de energía lumínica más pequeña posible, puede generar un potencial de receptor de 1 mV más o menos medible en un bastón. Ya con 30 fotones de luz se logrará producir la mitad de la saturación del bastón. ¿Cómo es posible que una cantidad tan pequeña de luz suscite una excitación tan potente? La respuesta señala que los fotorreceptores poseen una cascada química sumamente sensible que amplifica los efectos estimuladores en torno a un millón de veces, del modo siguiente:
1.El fotón activa un electrón en la porción 11-cis-retinal de la rodopsina; esto propicia la generación de metarrodopsina II, que es la forma activa de la rodopsina, 2. La rodopsina activada funciona como una enzima que estimula muchas moléculas de transducina, una proteína presente en su forma inactiva en las membranas de los discos y en la membrana celular del bastón.
3. La transducina activada estimula otras muchas moléculas más de fosfodiesterasa. 4. La fosfodiesterasa activada es otra enzima; su acción hidroliza de inmediato muchas moléculas de monofosfato de guanosina cíclico (GMPc), destruyéndolas. Antes de su desaparición, el GMPc se había ligado a la proteína del canal de sodio en la membrana externa del bastón de un modo que lo «inmoviliza» en su estado abierto. Pero ante la llegada de la luz, cuando la fosfodiesterasa hidroliza el GMPc, este hecho elimina la inmovilización y permite la oclusión de los canales de sodio. Por cada molécula de rodopsina activada inicialmente se cierran varios cientos de canales. Dado que el flujo de sodio a través de cada uno de estos canales ha sido rapidísimo, el paso de más de un millón de iones sodio queda bloqueado por el cierre del canal antes de que se abra de nuevo. Este descenso en la circulación del ion sodio es lo que excita al bastón, como ya se ha explicado.
5. En cuestión de 1 s más o menos, otra enzima, la cinasa de rodopsina, que está presente siempre en el bastón, desactiva la rodopsina activada (la metarrodopsina II), y toda la cascada se invierte hasta volver a su estado normal con los canales de sodio abiertos. Por tanto, los bastones han adquirido una importante cascada química que amplifica el efecto de un solo fotón de luz hasta provocar el movimiento de millones de iones sodio. Esto explica la sensibilidad extrema de estos receptores en condiciones de oscuridad. Los conos son de 30 a 300 veces menos sensibles que los bastones, pero incluso así permiten la visión de los colores a cualquier intensidad de luz superior a la de un crepúsculo sumamente tenebroso
