SENSIBILIDAD SOMÁTICAS: I. ORGANIZACIÓN GENERAL, LAS SENSACIONES TÁCTIL Y POSICIONAL
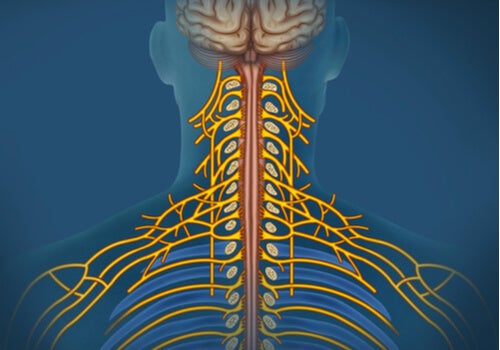
La sensibilidad somática es el mecanismo nervioso que recopila la información sensitiva de todo el cuerpo.
Las sensibilidades somáticas pueden clasificarse en tres tipos fisiológicos: 1) las sensibilidades somáticas mecanorreceptoras, formadas por las sensaciones táctiles y posicionales cuyo estímulo depende del desplazamiento mecánico de algún tejido del organismo; 2) las sensibilidades termorreceptoras, que detectan el calor y el frío, y 3) la sensibilidad al dolor, que se activa con factores que dañan los tejidos.
- La sensibilidad exterorreceptora es la que procede de la superficie del cuerpo.
- La sensibilidad propioceptiva es la que tiene que ver con el estado físico del cuerpo.
- La sensibilidad visceral es la que deriva de las vísceras del cuerpo.
- La sensibilidad profunda es la que viene de los tejidos profundos, como las fascias, los músculos y los huesos.

Caracteristicas de los receptores táctiles:
1 er lugar: algunas terminaciones nerviosas libres, que están distribuidas por todas partes en la piel y en otros muchos tejidos, son capaces de detectar el tacto y la presión.
2do lugar: un receptor al tacto dotado de una gran sensibilidad es el corpúsculo de Meissner, que es una terminación nerviosa encapsulada alargada perteneciente a una gran fibra nerviosa sensitiva mielínica (de tipo Aβ).
3er lugar: las yemas de los dedos y otras zonas que contienen una gran cantidad de
corpúsculos de Meissner también suelen albergar un número elevado de receptores táctiles de terminación bulbar.

4to lugar: el leve movimiento de cualquier pelo sobre el cuerpo estimula una fibra nerviosa que se enrosca en su base.
5to lugar: ubicados en las capas más profundas de la piel y también en los tejidos internos aún más profundos, hay muchas terminaciones de Ruffini, que son terminaciones encapsuladas multirramificadas.
6to lugar: los corpúsculos de Pacini, se hallan inmediatamente por debajo de la piel y quedan profundos en los tejidos de las fascias del organismo.

Casi todos los receptores sensitivos especializados, como los corpúsculos de Meissner, los receptores en cúpula de Iggo, los receptores pilosos, los corpúsculos de Pacini y las terminaciones de Ruffini, envían sus señales por fibras nerviosas de tipo Aβ que poseen una velocidad de transmisión entre 30 y 70 m/s. Por el contrario, los receptores táctiles de las terminaciones nerviosas libres mandan sus señales sobre todo a través de pequeñas fibras mielínicas de tipo Aδ que no conducen más que a una velocidad de 5 a 30 m/s.
Todos los receptores táctiles participan en la detección de las vibraciones, los corpúsculos de Pacini pueden identificar vibraciones con señales desde 30 hasta 800 ciclos/s debido a que responden con una rapidez extrema a las deformaciones minúsculas y veloces de los tejidos. También envían sus señales a las fibras nerviosas de tipo Aβ, que son capaces de transmitir hasta 1.000 impulsos por segundo.

Los estudios neurofisiológicos han demostrado la existencia de terminaciones nerviosas libres mecanorreceptoras de adaptación rápida y muy sensibles que tan solo suscitan sensaciones de cosquilleo y de picor. Estas terminaciones se encuentran casi exclusivamente en las capas superficiales de la piel, que también es el único tejido desde el que suelen poder despertarse estos fenómenos. Dichas sensaciones se transmiten por fibras amielínicas muy pequeñas de tipo C semejantes a las que se encargan del dolor de tipo lento y continuo.
Casi toda la información sensitiva procedente de los segmentos somáticos corporales penetra en la médula espinal a través de las raíces dorsales de los nervios raquídeos, dos vías sensitivas alternativas siguientes: 1) el sistema de la columna dorsal-lemnisco medial, o 2) el sistema anterolateral.
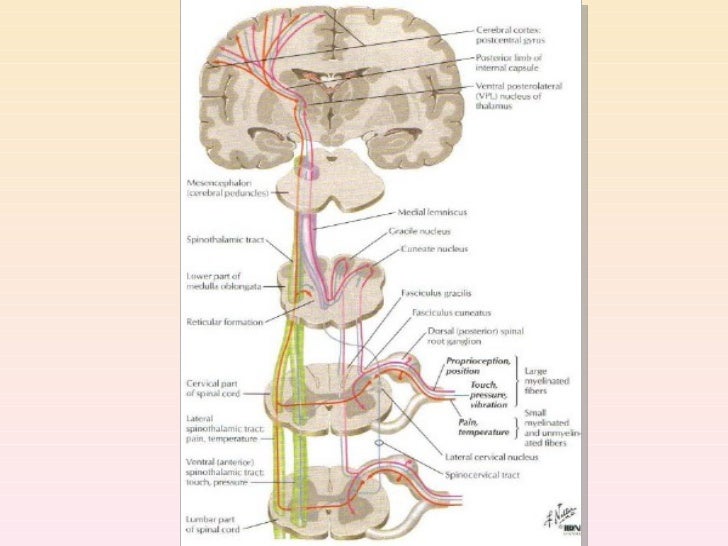
El sistema de la columna dorsal-lemnisco medial, como su nombre da a entender, transporta señales en sentido ascendente básicamente por las columnas dorsales de la médula hacia el bulbo raquídeo en el encéfalo. A continuación, después de hacer sinapsis y cruzar al lado opuesto a este nivel, siguen subiendo a través del tronco del encéfalo hasta el tálamo dentro del lemnisco medial. está compuesto por fibras nerviosas mielínicas grandes que transmiten señales hacia el cerebro a una velocidad de 30 a 110 m/s, mientras que el sistema anterolateral está integrado por fibras mielínicas más pequeñas cuya velocidad de transmisión oscila desde unos pocos metros por segundo hasta 40 m/s.
El sistema anterolateral posee una capacidad especial de la que carece el sistema dorsal, que es la propiedad de transmitir un amplio espectro de modalidades sensitivas como dolor, calor, frío y sensaciones táctiles groseras.

Al penetrar en la médula espinal a través de las raíces dorsales de los nervios raquídeos, las grandes fibras mielínicas procedentes de los mecanorreceptores especializados se dividen casi de inmediato para dar lugar a una rama medial y una rama lateral.
Las neuronas locales cumplen a su vez tres funciones:
- Una parte fundamental de ellas emite fibras que entran en las columnas dorsales de la médula y después ascienden hacia el encéfalo.
- Muchas de las fibras son muy cortas y acaban a nivel local en la sustancia gris de la médula espinal para producir los reflejos locales de esta estructura.
- Otras dan origen a los fascículos espinocerebelosos

Uno de los rasgos diferenciadores del sistema de la columna dorsal-lemnisco medial es la diversaorientación espacial que adquieren las fibras nerviosas procedentes de cada parte del cuerpo, y que se mantiene todo el tiempo,
La corteza cerebral humana, que manifiesta su división en unas 50 zonas distintas llamadas áreas de Brodmann según su diferente estructura histológica.

Las señales sensitivas pertenecientes a cualquier modalidad de sensación terminan en la corteza cerebral inmediatamente por detrás de la cisura central. También a grandes rasgos, la mitad anterior del lóbulo parietal se ocupa casi por completo de la recepción e interpretación de las señales somatosensitivas, pero la mitad posterior aporta unos niveles aún más altos de interpretación.
Las señales visuales acaban en el lóbulo occipital, y las señales auditivas terminan en el lóbulo temporal.
Áreas sensitivas independientes en el lóbulo parietal anterior, llamadas área somatosensitiva I y área somatosensitiva II.
El área somatosensitiva I es mucho más extensa e importante que el área somatosensitiva II, hasta el punto de que, en el uso popular, el término «corteza somatosensitiva» casi siempre significa área I.

El área somatosensitiva I se halla inmediatamente detrás de la cisura central, situada en la circunvolución poscentral de la corteza cerebral humana.
La corteza cerebral contiene seis capas de neuronas, comenzando por la capa I próxima a la superficie cerebral y siguiendo cada vez por zonas más profundas hasta la capa VI
- La señal sensitiva entrante excita en primer lugar la capa neuronal IV; a continuación, se propaga hacia la superficie de la corteza y también hacia otras capas más profundas.
- Las capas I y II reciben señales de entrada difusas inespecíficas procedentes de los centros inferiores del encéfalo, que facilitan regiones corticales específicas.
- Las neuronas de las capas II y III envían axones hacia las porciones emparentadas entre sí de la corteza cerebral en el lado opuesto del cerebro a través del cuerpo calloso.
- Las neuronas de las capas V y VI mandan axones hacia las partes más profundas del sistema nervioso.
La corteza sensitiva está organizada en columnas verticales de neuronas; cada columna detecta un lugar sensitivo diferente en el cuerpo con una modalidad sensitiva específica, las neuronas de la corteza somatosensitiva están dispuestas formando columnas verticales que se extienden a lo largo de las seis capas corticales, con un diámetro de 0,3 a 0,5 mm y un contenido quizá de 10.000 somas neuronales.
Las áreas 5 y 7 de Brodmann de la corteza cerebral, situadas en la corteza parietal detrás del área
somatosensitiva I, ocupan un lugar importante en la labor de descifrar los significados más profundos de la información sensitiva en las áreas somatosensitivas. Por tanto, se las denomina áreas de asociación somatosensitiva.
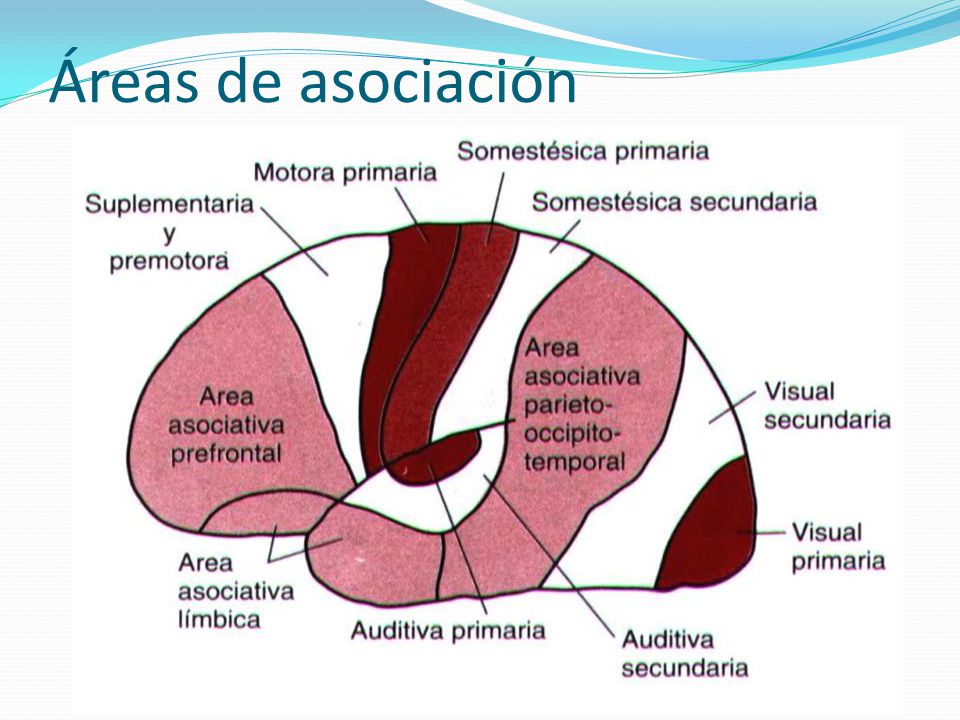
Esta situación también encaja con la disposición anatómica de los fascículos neuronales que penetran en el área de asociación somatosensitiva, porque recibe señales desde: 1) el área somatosensitiva I; 2) los núcleos ventrobasales del tálamo; 3) otras zonas talámicas; 4) la corteza visual, y 5) la corteza auditiva.
Un estímulo débil solo causa el disparo de las neuronas más centrales. Otro más intenso provoca el disparo de más neuronas aún, pero las del centro descargan a una frecuencia considerablemente superior que las que se encuentran más alejadas.

Un método empleado a menudo para verificar la propiedad táctil de la distinción consiste en determinar la denominada capacidad discriminatoria entre «dos puntos» de una persona. En esta prueba se presiona suavemente la piel con dos agujas al mismo tiempo y la persona señala si siente el estímulo de uno o de dos puntos.

Las sensibilidades posicionales también se denominan a menudo sensibilidades propioceptivas y pueden dividirse en dos subtipos: 1) sensibilidad posicional estática, que significa la percepción consciente de la orientación de las diferentes partes del cuerpo unas respecto a otras, y 2) velocidad de la sensibilidad al movimiento, también llamada cinestesia o propiocepción dinámica.
El conocimiento de la posición, tanto estática como dinámica, depende de la información sobre el grado de angulación de todas las articulaciones en cualquiera de los planos y sus velocidades de cambio. Por tanto, son múltiples los diferentes tipos de receptores que sirven para determinar la angulación articular y que se emplean en conjunto dentro de la sensibilidad posicional.
Entre los receptores más relevantes que sirven para determinar la angulación articular en el recorrido medio del movimiento figuran los husos musculares.

Los corpúsculos de Pacini y los husos musculares están especialmente adaptados para detectar una velocidad de cambio rápida. Es probable que se trate de los receptores con una mayor responsabilidad de averiguar la velocidad del movimiento.
La vía anterolateral, encargada de la transmisión de señales sensitivas ascendentes por la médula espinal y en dirección al encéfalo, al revés que la vía de la columna dorsal, transporta unos tipos que no requieren una localización muy diferenciada de la fuente de origen ni tampoco una distinción en cuanto a las gradaciones finas de intensidad.
Los mismos principios que sirven para el sistema de la columna dorsal-lemnisco medial se aplican a la transmisión por la vía anterolateral, exceptuando las siguientes diferencias: 1) la velocidad de transmisión solo llega a un tercio o la mitad de la que posee el sistema de la columna dorsal- lemnisco medial, y oscila entre 8 y 40 m/s; 2) el grado de localización espacial de las señales es escaso; 3) la gradación de las intensidades también es mucho menos precisa, y en la mayoría de las sensaciones se identifican de 10 a 20, en vez de alcanzar las 100 como el sistema de la columna dorsal, y 4) la capacidad para transmitir señales que se repitan o varíen con rapidez es mala.
SENSIBILIDADES SOMÁTICAS: II. DOLOR CEFALEA Y SENSIBILIDAD TÉRMICA

El dolor aparece siempre que cualquier tejido resulta dañado y hace que el individuo reaccione apartando el estímulo doloroso.
El dolor se ha clasificado en dos tipos fundamentales: dolor rápido y dolor lento. El dolor rápido se siente en cuestión de 0,1 s después de haber aplicado el estímulo correspondiente, mientras que el dolor lento no empieza hasta pasado un mínimo de 1 s y a continuación crece con lentitud a lo largo de muchos segundos y en ocasiones hasta minutos.
El dolor rápido también se describe con otros muchos nombres alternativos, como dolor intenso, dolor punzante, dolor agudo y dolor eléctrico. Este tipo de dolor se siente cuando se clava una aguja en la piel, cuando se corta con un cuchillo o cuando sufre una quemadura intensa. El dolor rápido y agudo no se siente en los tejidos más profundos del organismo.
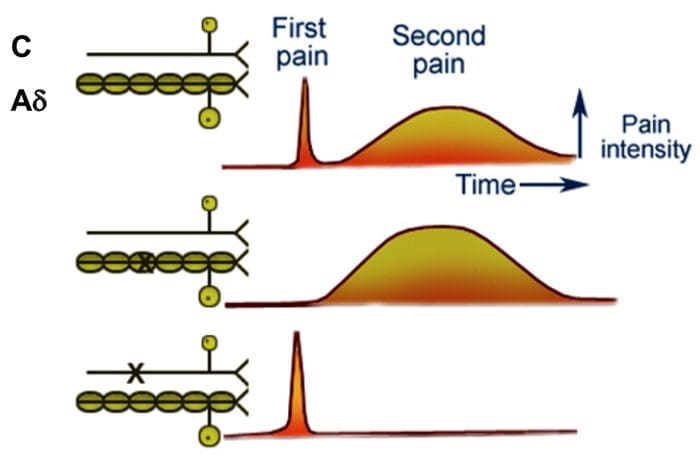
El dolor lento también se designa con muchos nombres, entre ellos dolor lento urente, dolor sordo, dolor pulsátil, dolor nauseoso y dolor crónico. Este tipo de dolor suele ir asociado a una destrucción tisular.
Los receptores para el dolor de la piel y de otros tejidos siempre son terminaciones nerviosas libres. Se encuentran extendidos por las capas superficiales de la piel así como en ciertos tejidos internos, como el periostio, las paredes arteriales, las superficies articulares, y las hoces y la tienda en la bóveda craneal.
El dolor puede despertarse por múltiples tipos de estímulos, que se dividen en estímulos dolorosos mecánicos, térmicos y químicos. A grandes rasgos, el dolor rápido se suscita a partir de los tipos de estímulo mecánico y térmico, mientras que el dolor lento puede surgir con cualquiera de los tres.

Algunos de los productos que excitan el dolor de tipo químico son bradicinina, serotonina, histamina, iones potasio, ácidos, acetilcolina y enzimas proteolíticas. Además, las prostaglandinas y la sustancia P favorecen la sensibilidad de las terminaciones para el dolor, pero no las activan directamente.
La adaptación de los receptores para el dolor es muy escasa y a veces nula en absoluto. En realidad, bajo ciertas condiciones, la excitación de las fibras para el dolor crece cada vez más, sobre todo en el tipo lento, sordo, nauseoso,si el estímulo doloroso persiste. Este aumento de la sensibilidad en los receptores para el dolor se llama hiperalgesia.
La intensidad del dolor también mantiene una fiel relación con la velocidad de la lesión tisular ocasionada por otras fuentes aparte del calor, como las infecciones bacterianas, la isquemia del tejido, una contusión tisular.

Los extractos de tejido dañado provocan un dolor intenso cuando se inyectan bajo la piel normal. Una sustancia que parece más dolorosa que las demás es la bradicinina. Los investigadores han propuesto que este podría ser el agente con una mayor responsabilidad sobre el dolor generado después de un daño tisular.
Cuando queda bloqueado el flujo sanguíneo hacia un tejido, este suele volverse muy doloroso en cuestión de unos minutos.
Una de las causas propuestas para explicar el dolor que existe durante la isquemia es la acumulación de grandes cantidades de ácido láctico en los tejidos, surgido a raíz del metabolismo anaerobio (es decir, metabolismo sin oxígeno).

El espasmo muscular también es una causa frecuente de dolor y representa el fundamento de muchos síndromes clínicos dolorosos.
El espasmo acelera el metabolismo del tejido muscular, lo que acentúa aún más la isquemia relativa, y crea las condiciones ideales para la liberación de sustancias químicas inductoras de dolor.
A pesar de que todos los receptores para el dolor consisten en terminaciones nerviosas libres, estas estructuras utilizan dos vías distintas para transmitir sus señales respectivas hacia el sistema nervioso central. Ambas guardan una correspondencia básica con los dos tipos de dolor: una vía para el dolor rápido agudo y otra vía para el dolor lento crónico.
Las señales correspondientes al dolor rápido agudo nacen con estímulos dolorosos de tipo mecánico o térmico. Su transmisión sigue los nervios periféricos hasta la médula espinal a través de pequeñas fibras de tipo Aδ a una velocidad entre 6 y 30 m/s. Por el contrario, el dolor de carácter lento crónico se suscita sobre todo a partir de los estímulos químicos correspondientes, pero a veces también con estímulos mecánicos o térmicos persistentes. Este dolor lento crónico llega a la médula espinal por medio de las fibras de tipo C a una velocidad entre 0,5 y 2 m/s.

El dolor agudo informa a gran velocidad sobre la situación lesiva y, por tanto, cumple una función importante para conseguir que la persona reaccione de inmediato y se aparte del estímulo. El dolor lento tiende a crecer con el tiempo. Esta sensación produce a la larga el dolor intolerable y obliga a que la persona trate de mitigar su causa.
En la médula espinal,las señales de dolor toman dos caminos hacia el encéfalo, a través: 1) del fascículo neoespinotalámico, y 2) del fascículo paleoespinotalámico.en la médula espinallas señales de dolor toman dos caminos hacia el encéfalo, a través: 1) del fascículo neoespinotalámico, y 2) del fascículo paleoespinotalámico. Fascículoneoespinotalámico para el dolor rápido. Las fibras rápidas para el dolor de tipo Aδ transmiten básicamente esta sensación en la modalidad térmica aguda y mecánica. Acaban sobre todo en la lámina I (lámina marginal) de las astas dorsales, y allí excitan las neuronas de segundo orden pertenecientes al fascículo neoespinotalámico.

Terminación del fascículo neoespinotalámico en el tronco del encéfalo y el tálamo. Unas pocas fibras del fascículo neoespinotalámico acaban en la formación reticular del tronco del encéfalo, pero la mayoría pasan de largo hacia el tálamo sin realizar paradas, y terminan en el complejo ventrobasal junto al fascículo de la columna dorsal-lemnisco medial encargado de la sensibilidad táctil.
Capacidad del sistema nervioso para localizar el dolor rápido en el cuerpo. El dolor de tipo rápido agudo puede localizarse con mucha mayor precisión en las diversas partes del cuerpo que el dolor lento crónico. Sin embargo, cuando no se estimulan más que los receptores para el dolor, sin una activación simultánea de los receptores táctiles, incluso el dolor rápido puede estar poco localizado, muchas veces solo con un margen de unos 10 cm alrededor de la zona en cuestión.
Glutamato, el neurotransmisor más probable de las fibras para el dolor rápido de tipo
Aδ. Se piensa que el glutamato es la sustancia neurotransmisora segregada en la médula espinal por las terminaciones de las fibras nerviosas para el dolor de tipo Aδ. El glutamato es uno de los transmisores excitadores que recibe un uso más generalizado en el sistema nervioso central, cuya acción normalmente no dura nada más que unos pocos milisegundos.
Vía paleoespinotalámica para la transmisión del dolor lento crónico. La vía paleoespinotalámica es un sistema mucho más antiguo y básicamente transmite el dolor procedente de las fibras periféricas de tipo C dotado de un carácter lento crónico, aunque también transporta algunas señales correspondientes a las fibras de tipo Aδ. En esta vía, dichas fibras periféricas acaban en la médula espinal casi en su integridad entre las láminas II y III de las astas dorsales, que en conjunto reciben el nombre de sustancia gelatinosa.
Sustancia P, el neurotransmisor más probable de las terminaciones nerviosas con un carácter lento crónico de tipo C. Las investigaciones señalan que los terminales de las fibras para el dolor de tipo C que entran en la médula espinal segregan los transmisores glutamato y sustancia P. El glutamato actúa de manera inmediata y solo dura unos pocos milisegundos. La sustancia P se libera con mucha mayor lentitud, acumulándose su concentración durante un período de segundos o incluso de minutos.
La vía paleoespinotalámica lenta crónica presenta un final amplio en el tronco del encéfalo, la mayoría acaban en una de las tres áreas siguientes: 1) los núcleos de la formación reticular del bulbo raquídeo, la protuberancia y el mesencéfalo; 2) la región tectal del mesencéfalo profunda a los colículos superiores e inferiores, o 3) la zona gris periacueductal que rodea al acueducto de Silvio.
La localización del dolor transmitido a través de la vía paleoespinotalámica es imprecisa. La resección íntegra de las áreas sensitivas somáticas de la corteza cerebral no evita la percepción del dolor.
La estimulación eléctrica de las regiones reticulares del tronco del encéfalo y de los núcleos intralaminares del tálamo, las zonas donde acaba el dolor de tipo lento que causa sufrimiento, posee un potente efecto potenciador de la actividad nerviosa por todo el encéfalo.
La cordotomía no siempre tiene éxito para calmar el dolor, por dos razones. En primer lugar, muchas fibras que transportan esta sensación y proceden de la parte superior del cuerpo no cruzan hacia el lado opuesto de la médula espinal hasta después de haber llegado al encéfalo, por lo que la cordotomía no las divide. En segundo lugar, el dolor suele volver varios meses más tarde, en parte como consecuencia de la sensibilización experimentada por otras vías que en condiciones normales son demasiado tenues como para resultar eficaces.

El grado con el que cada persona reacciona frente al dolor varía tremendamente. Esta variación obedece en parte a una propiedad que posee el encéfalo en sí mismo para suprimir la entrada de señales dolorosas al sistema nervioso mediante la activación de un mecanismo para controlar el dolor, llamado sistema de analgesia.
Consta de tres componentes fundamentales: (1) la región gris periacueductal y las áreas periventriculares del mesencéfalo y la parte superior de la protuberancia que rodean al acueducto de Silvio y a las porciones del tercer y del cuarto ventrículos, (2) el núcleo magno del rafe, un núcleo delgado de la línea media situado en las partes inferior de la protuberancia y superior del bulbo raquídeo, y el núcleo reticular paragigantocelular, que ocupa una posición lateral en este último y la médula espinal hacia (3) un complejo inhibidor del dolor localizado en las astas dorsales de la médula espinal.

La estimulación eléctrica de la región gris periacueductal o del núcleo magno del rafe es capaz de suprimir muchas señales de dolor potentes que penetran a través de las raíces medulares dorsales.
Diversas sustancias transmisoras participan en el sistema analgésico, especialmente la encefalina y la serotonina. Se cree que la encefalina propicia una inhibición presináptica y postsináptica de las fibras para el dolor de tipo C y Aδ al hacer sinapsis en las astas dorsales.
Productos de degradación de tres grandes moléculas proteicas: proopiomelanocortina, proencefalina y prodinorfina. Entre los más importantes de estos compuestos figuran la β-endorfina, la metencefalina, la leuencefalina y la dinorfina.
Muchas veces una persona siente dolor en una parte del cuerpo situada bastante alejada del tejido que lo origina. Este fenómeno se llama dolor referido.
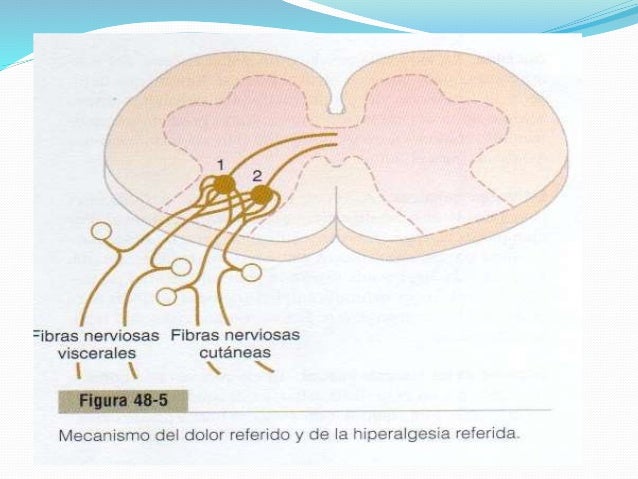
Cuando se estimulan estas fibras, las señales de dolor procedentes de las vísceras viajan al menos a través de algunas de las mismas neuronas que conducen esta información desde la piel, y la persona recibe la percepción de que las sensaciones se originan en la piel.
El dolor procedente de las diferentes vísceras del abdomen y del tórax es uno de los escasos criterios que pueden utilizarse para diagnosticar una inflamación visceral, las enfermedades infecciosas y otros padecimientos a este nivel.
Cualquier fenómeno que produzca una estimulación difusa de las terminaciones nerviosas para el dolor en una víscera provoca un dolor intenso. Cualquier estímulo que excite las terminaciones nerviosas para el dolor en regiones difusas de las vísceras puede suscitar un dolor visceral.
Cuando una enfermedad afecta a una víscera, el proceso patológico a menudo se propaga al peritoneo parietal, la pleura o el pericardio. Estas superficies parietales, lo mismo que la piel, reciben una amplia inervación dolorosa de los nervios raquídeos periféricos.
El ser humano puede percibir diferentes gradaciones de frío y de calor, desde ambientes helados a fríos, frescos, indiferentes, templados, cálidos o ardientes.
Las gradaciones térmicas se distinguen como mínimo por tres tipos de receptores sensitivos: receptores para el frío, receptores para el calor y receptores para el dolor. Los receptores para el dolor se estimulan únicamente ante un grado extremo de calor o de frío y, por tanto, son responsables, junto los receptores para el frío y para el calor, de las sensaciones de «frío helado» y «calor ardiente».
