RECEPTORES SENSITIVOS, CIRCUITOS NEURONALES PARA EL PROCESAMIENTO DE LA INFORMACIÓN.

Nuestras percepciones sobre las señales del cuerpo y del mundo que nos rodea están mediadas por un complejo sistema de receptores sensitivos que detectan estímulos como el tacto, el sonido, la luz, el dolor, el frío y el calor.
Clasifica cinco tipos básicos de receptores sensitivos: 1) mecanorreceptores, que detectan la compresión mecánica o su estiramiento, o el de los tejidos adyacentes; 2) termorreceptores, que detectan los cambios en la temperatura, donde algunos de los receptores se encargan del frío y otros del calor; 3) nocirreceptores (receptores del dolor), que detectan daños físicos o químicos que se producen en los tejidos; 4) receptores electromagnéticos, que detectan la luz en la retina ocular, y 5) quimiorreceptores, que detectan el gusto en la boca, el olfato en la nariz, la cantidad de oxígeno en la sangre arterial, la osmolalidad de los líquidos corporales, la concentración de dióxido de carbono y otros factores que completen la bioquímica del organismo.

Cada uno de los principales tipos sensitivos que podemos experimentar, dolor, tacto, visión, sonido, etc., se llama modalidad de sensación. La respuesta señala que cada fascículo nervioso termina en un punto específico del sistema nervioso central y el tipo de sensación vivida cuando se estimula una fibra nerviosa queda determinado por la zona del sistema nervioso a la que conduce esta fibra.
Las fibras nerviosas para transmitir nada más que una modalidad de sensación se llama principio de la línea marcada.

Todos los receptores sensitivos tienen un rasgo en común. Cualquiera que sea el tipo de estímulo que les excite, su efecto inmediato consiste en modificar su potencial eléctrico de membrana.
Los diversos receptores pueden excitarse siguiendo alguno de los siguientes modos de generar potenciales de receptor: 1) por deformación mecánica del receptor, que estire su membrana y abra los canales iónicos; 2) por la aplicación de un producto químico a la membrana, que también abra los canales iónicos; 3) por un cambio de la temperatura de la membrana, que modifique su permeabilidad, o 4) por los efectos de la radiación electromagnética, como la luz que incide sobre un receptor visual de la retina, al modificar directa o indirectamente las características de la membrana del receptor y permitir el flujo de iones a través de sus canales.
La amplitud máxima de la mayoría de los potenciales de receptor sensitivos es de unos 100 mV, pero este valor no se alcanza más que cuando la intensidad del estímulo correspondiente es altísima.

El corpúsculo de Pacini posee una fibra nerviosa central que recorre su núcleo. Alrededor de esta fibra nerviosa central hay una cápsula compuesta por múltiples capas concéntricas, de manera que la compresión del corpúsculo desde fuera sobre cualquier punto alargará, oprimirá o deformará la fibra central de cualquier otro modo.

La frecuencia de los potenciales de acción repetidos que se transmiten desde los receptores sensitivos aumenta de forma aproximadamente proporcional al incremento del potencial de receptor.
Se trata de un principio sumamente importante que resulta aplicable a casi todos los receptores sensitivos; les dota de sensibilidad frente a experiencias sensitivas muy débiles, sin llegar a una frecuencia máxima de disparo hasta que la experiencia sea de tremenda magnitud. Esta característica ofrece al receptor una enorme gama de respuesta, desde un extremo muy débil hasta otro muy intenso.

Los receptores sensitivos es su adaptación parcial o total a cualquier estímulo constante después de haber transcurrido un tiempo. Es decir, cuando se aplica un estímulo sensitivo continuo, el receptor responde al principio con una frecuencia de impulsos alta y después baja cada vez más hasta que acaba disminuyendo la frecuencia de los potenciales de acción para pasar a ser muy pocos o muchas veces desaparecer del todo.

La capacidad de adaptación de ciertos receptores sensitivos es mucho mayor que la de
otros.
El mecanismo de adaptación varía con cada tipo de receptor, básicamente lo mismo que la producción de un potencial de receptor constituye una propiedad individual.
Los receptores de adaptación lenta siguen transmitiendo impulsos hacia el cerebro mientras siga presente el estímulo. Mantienen al cerebro constantemente informado sobre la situación del cuerpo y su relación con el medio.
Otros receptores de adaptación lenta son los siguientes: 1) los pertenecientes a la mácula en el aparato vestibular; 2) los receptores para el dolor; 3) los barorreceptores del árbol arterial, y 4) los quimiorreceptores de los cuerpos carotídeo y aórtico.
Los receptores que se adaptan con rapidez no pueden utilizarse para transmitir una señal continua debido a que solo se activan cuando cambia la intensidad del estímulo. Con todo, reaccionan potentemente siempre que esté teniendo lugar un ambio de hecho. Por tanto, se llaman receptores de velocidad, receptores de movimiento o receptores fásicos.
Si se conoce la velocidad a la que tiene lugar un cambio en la situación corporal, se podrá predecir cuál será el estado del organismo a su juicio unos cuantos segundos o incluso minutos más tarde.
Algunas señales necesitan transmitirse con enorme rapidez hacia el sistema nervioso central o salir de él; si no, la información resultaría inútil. La gama de las velocidades de conducción oscila entre 0,5 y 120 m/s.
La sumación espacial, por el cual se transmite la intensidad creciente de una señal mediante un número progresivamente mayor de fibras.

Cada una de estas fibras se ramifica en cientos de minúsculas terminaciones nerviosas libres que sirven como receptores para el dolor.
Un segundo medio para transmitir señales de intensidad creciente consiste en acelerar la frecuencia de los impulsos nerviosos que recorren cada fibra, lo que se denomina sumación temporal.
El sistema nervioso central está integrado por miles de millones de grupos neuronales; algunos de estos grupos contienen unas cuantas neuronas, mientras que otros presentan una cantidad enorme.
Cada grupo neuronal posee su propia organización especial que le hace procesar las señales de un modo particular y singular, lo que permite que el agregado total de grupos cumpla la multitud de funciones del sistema nervioso.

Cada fibra que llega se divide cientos o miles de veces, aportando mil fibrillas terminales como mínimo que se esparcen por una gran zona dentro del grupo para hacer sinapsis con las dendritas o los somas de sus neuronas. La zona neuronal estimulada por cada fibra nerviosa que entra se llama campo de estimulación.
Una versión muy condensada de un grupo neuronal porque cada fibra nerviosa de entrada suele suministrar un enorme número de terminales ramificados a los cientos o miles de neuronas situadas en su «campo» de distribución.
En la porción central del campo representado en esta última imagen, indicada por el área contenido dentro del círculo, todas las neuronas están estimuladas por la fibra que llega. Por tanto, se dice que esta es la zona de descarga de la fibra de entrada, también llamada zona excitada o zona liminal. A cada lado, las neuronas están facilitadas pero no excitadas, y estas áreas se llaman zona facilitada, o también zona por debajo del umbral o zona subliminal.

Algunas fibras de entrada inhiben a las neuronas, en vez de excitarlas. Este mecanismo es el opuesto a la facilitación, y el campo de las ramas inhibidoras en su integridad se llama zona inhibidora.
Las señales débiles que penetran en un grupo neuronal acaben excitando a una cantidad mucho mayor de las fibras nerviosas que lo abandonan. Este fenómeno se llama divergencia.
Divergencia amplificador, esta divergencia significa sencillamente que una señal de entrada se disemina sobre un número creciente de neuronas a medida que atraviesa sucesivos órdenes de células en su camino, es el tipo de divergencia característico de la
vía corticoespinal.
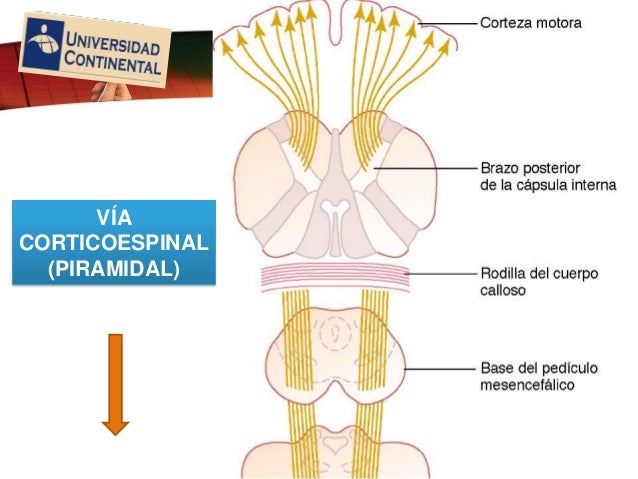
Divergencia en múltiples fascículos

La convergencia significa que un conjunto de señales procedentes de múltiples orígenes se reúnen para excitar una neurona concreta.
La convergencia desde una sola fuente, es decir, numerosos terminales derivados de la llegada de un solo fascículo de fibras acaban en lamisma neurona.

Los potenciales de acción que convergen sobre la neurona desde muchos terminales proporcionan una sumación espacial suficiente para llevar a la célula hasta el umbral necesario de descarga.
La convergencia también puede surgir con las señales de entrada (excitadoras o inhibidoras) derivadas de múltiples fuentes.
Esta convergencia permite la sumación de información derivada de diversas fuentes y la respuesta resultante reúne el efecto acumulado de todos los diferentes tipos de información. La convergencia es uno de los medios importantes que utiliza el sistema nervioso central para relacionar, sumar y clasificar distintas clases de información.

La fibra de entrada activa directamente la vía de salida excitadora, pero estimula una neurona inhibidora intermedia (neurona 2), que segrega un tipo diferente de sustancia transmisora encargada de inhibir la segunda vía de salida desde el grupo. Esta clase de circuito también resulta importante para evitar la hiperactividad en muchas porciones del cerebro.
Una señal que penetra en un grupo suscita una descarga de salida prolongada, llamada posdescarga, cuya duración va desde unos pocos milisegundos hasta muchos minutos después de que haya acabado la señal de entrada.
Uno de los circuitos más importantes del sistema nervioso es el circuito reverberante u oscilatorio.
Está ocasionado por una retroalimentación positiva dentro del circuito neuronal que ejerce una retroalimentación encargada de reexcitar la entrada del mismo circuito.

Algunos circuitos neuronales emiten señales de salida de forma continua, incluso sin señales deentrada excitadoras. Al menos dos mecanismos pueden ocasionar este efecto: 1) la descarga neuronal intrínseca continua, y 2) las señales reverberantes continuas.
