SISTEMA NERVIOSO: PRINCIPIOS GENERALES Y FISIOLOGÍA DE LA SENSIBILIDAD.
ORGANIZACIÓN DEL SISTEMA NERVIOSO, FUNCIONES BÁSICAS DE LAS SINAPSIS Y NEUROTRANSMISORES
El sistema nervioso central contiene más de 100.000 millones de neuronas. Las señales de entrada llegan a ella a través de las sinapsis situadas fundamentalmente en las dendritas neuronales, pero también en el soma celular. Las conexiones sinápticas procedentes de las fibras aferentes pueden ser tan solo unos cientos o llegar hasta 200,000. Un rasgo especial de la mayoría de las sinapsis consiste en que normalmente la señal sólo circula en sentido anterógrado.
La mayoría de las actividades del sistema nervioso se ponen en marcha cuando las experiencias sensitivas excitan los receptores sensitivos, ya sean de carácter visual en los ojos, auditivo en los oídos, táctil en la superficie del organismo o de otros tipos.
Esta información penetra en el sistema nervioso central a través de los nervios periféricos y se transporta de inmediato hasta múltiples zonas sensitivas en: 1) la médula espinal a todos sus niveles; 2) la formación reticular del bulbo raquídeo, la protuberancia y el mesencéfalo en el encéfalo; 3) el cerebelo; 4) el tálamo, y 5) áreas de la corteza cerebral.

La misión más importante del sistema nervioso consiste en regular las diversas actividades del organismo. Para desempeñar esta tarea, debe controlar los siguientes aspectos: 1) la contracción de los músculos esqueléticos adecuados en todo el cuerpo; 2) la contracción de la musculatura lisa de las vísceras, y 3) la secreción de sustancias químicas activas por parte de las glándulas exocrinas y endocrinas en muchas zonas del organismo, denominan funciones motoras del sistema nervioso.

El eje nervioso motor «esquelético» del sistema nervioso cuya actividad está dedicada a controlar la contracción de la musculatura esquelética.
Una de las funciones más importantes del sistema nervioso consiste en elaborar la información que le llega de tal modo que dé lugar a las respuestas motoras y mentales adecuadas. El encéfalo descarta más del 99% de toda la información sensitiva que recibe por carecer de interés o de importancia.
Cuando una información sensitiva importante excita la mente, de inmediato resulta encauzada hacia las regiones motoras e integradoras oportunas del encéfalo para suscitar las respuestas deseadas. Esta canalización y tratamiento de la información se denomina función integradora del sistema nervioso.
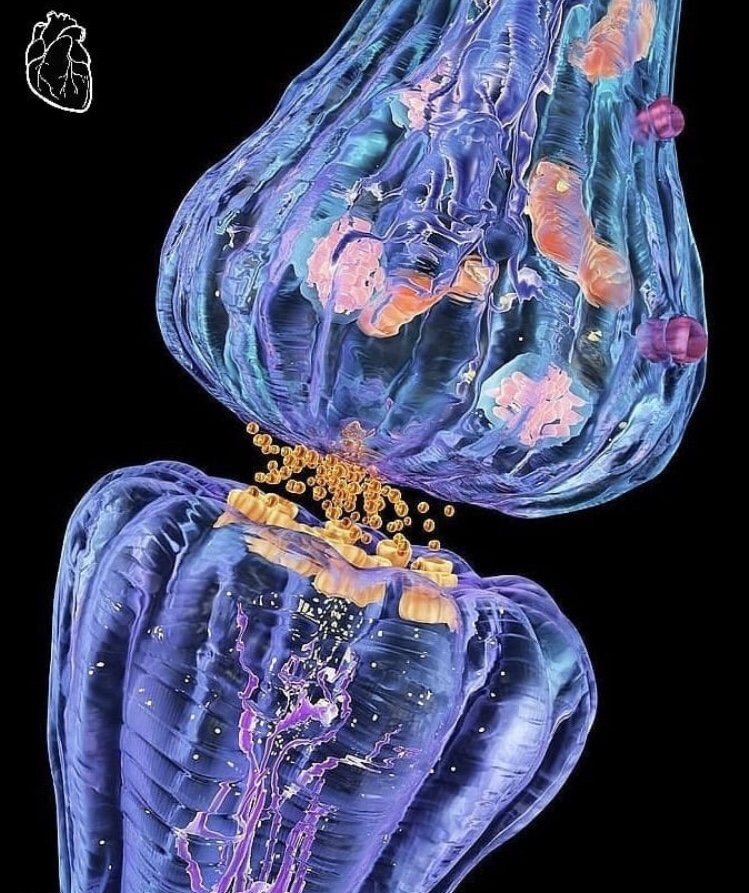
La sinapsis es el punto de unión de una neurona con la siguiente, determinan las direcciones de propagación que toma cualquier señal por el sistema nervioso. Las señales facilitadoras e inhibidoras procedentes de otras regiones del sistema nervioso tienen la capacidad de controlar la transmisión sináptica, a veces abriendo las sinapsis para efectuar la comunicación y en otras ocasiones cerrándolas.
La acumulación de la información es el proceso que llamamos memoria, y también constituye una función de las sinapsis. Cada vez que determinados tipos de señales sensitivas atraviesan una secuencia de sinapsis, estas adquieren una mayor capacidad para transmitir ese mismo tipo de señal la próxima vez, situación que llamamos facilitación.
Una vez que los recuerdos están guardados en el sistema nervioso, pasan a formar parte de los mecanismos de procesamiento cerebral para el «pensamiento» en el futuro. Es decir, los procedimientos de deliberación del encéfalo comparan las experiencias sensitivas nuevas con los recuerdos acumulados.

El sistema nervioso humano ha heredado unas capacidades funcionales especiales correspondientes a cada etapa recorrida por el desarrollo evolutivo del hombre. A partir de este bagaje, los principales niveles del sistema nervioso central que presentan unas características funcionales específicas son tres: 1) el nivel medular; 2) el nivel encefálico inferior o subcortical, y 3) el nivel encefálico superior o cortical.

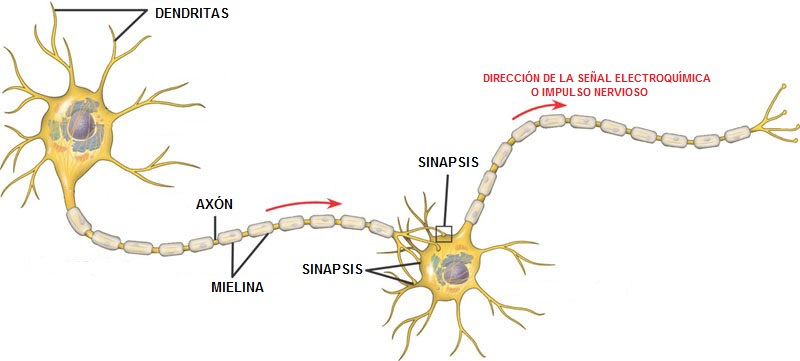
La información recorre el sistema nervioso central sobre todo bajo la forma de potenciales de acción nerviosos, llamados simplemente impulsos nerviosos, a través de una sucesión de neuronas, una después de la otra. Sin embargo, además, cada impulso puede: 1) quedar bloqueado en su transmisión de una neurona a la siguiente; 2) convertirse en una cadena repetitiva a partir de un solo impulso, o 3) integrarse con los procedentes de otras células para originar patrones muy intrincados en las neuronas sucesivas. Existen dos tipos de sinapsis: químicas y eléctricas.

La mayoría de las sinapsis utilizadas para la transmisión de señales en el sistema nervioso central del ser humano son sinapsis químicas. En estas sinapsis, la primera neurona segrega un producto químico denominado neurotransmisor (a menudo llamado sustancia transmisora) a nivel de la terminación nerviosa. Entre las mejor conocidas figuran las siguientes: acetilcolina, noradrenalina, adrenalina, histamina, ácido γ -aminobutírico (GABA), glicina, serotonina y glutamato.
En las sinapsis eléctricas los citoplasmas de las células adyacentes están conectados directamente por grupos de canales de iones llamados uniones en hendidura que permiten el movimiento libre de los iones desde el interior de una célula hasta el interior de la siguiente.
Aunque la mayoría de las sinapsis en el encéfalo son químicas, en el sistema nervioso central pueden coexistir sinapsis eléctricas y químicas. La transmisión bidireccional de las sinapsis eléctricas les permite colaborar en la coordinación de las actividades de grandes grupos de neuronas interconectadas.

Las sinapsis químicas poseen una característica sumamente importante que las convierte en un elemento muy conveniente para transmitir la mayor parte de las señales en el sistema nervioso. Esta característica hace posible que siempre conduzcan las señales en un solo sentido: es decir, desde la neurona que segrega el neurotransmisor, denominada neurona presináptica, hasta la neurona sobre la que actúa el transmisor, llamada neurona postsináptica.
La sinapsis está compuesta por tres partes fundamentales: el soma, que es el cuerpo principal de la neurona; el único axón, que se extiende desde el soma hacia un nervio periférico para abandonar la médula espinal, y las dendritas, que constituyen una gran cantidad de prolongaciones ramificadas del soma con unas dimensiones hasta de 1 mm de recorrido hacia las zonas adyacentes en la médula.

Las neuronas pertenecientes a otras porciones de la médula y el encéfalo se distinguen de la motoneurona anterior en los siguientes aspectos: 1) las dimensiones del soma celular; 2) la longitud, el tamaño y el número de dendritas, que oscila desde casi cero a muchos centímetros; 3) la longitud y el tamaño del axón, y 4) el número de terminales presinápticos, que puede oscilar desde tan solo unos
pocos hasta llegar a 200.000.
Las terminales presinápticas se parecen a pequeños botones redondos u ovalados; de ahí que a veces se les llame botones terminales, botones, pies terminales o botones sinápticos.Terminales presinápticosse parecen a pequeños botones redondos u ovalados; de ahí que a veces se les llame botones terminales, botones, pies terminales o botones sinápticos.
En él existen dos estructuras internas de importancia para la función excitadora o inhibidora de la sinapsis: las vesículas transmisoras y las mitocondrias. Las vesículas transmisoras contienen el neurotransmisor que, cuando se libera a la hendidura sináptica, excita o inhibe la neurona postsináptica. Las mitocondrias aportan trifosfato de adenosina (ATP), que a su vez suministra energía para sintetizar más sustancia transmisora.

La membrana de la neurona postsináptica contiene una gran cantidad de proteínas receptoras. Las moléculas de estos receptores poseen dos elementos importantes: 1) un componente de unión que sobresale fuera desde la membrana hacia la hendidura sináptica y donde se fija el neurotransmisor procedente del terminal presináptico, y 2) un componente intracelular que atraviesa toda la membrana postsináptica hasta el interior de la neurona postsináptica.
Los receptores de neurotransmisores que activan directamente los canales iónicos a menudo se denominan receptores ionotrópicos, mientras que los que actúan a través de sistemas de segundos mensajeros reciben el nombre de receptores metabotrópicos.
Los canales iónicos de la membrana neuronal postsináptica suelen ser de dos tipos: 1) canales catiónicos, cuya clase más frecuente deja pasar iones sodio cuando se abren, pero a veces también cumplen esta función con el potasio o el calcio, y 2) canales aniónicos, que permiten sobre todo el paso de los iones cloruro, pero también de minúsculas cantidades de otros aniones. Los canales catiónicos que transportan iones sodio están revestidos de cargas negativas.
Los canales iónicos no son idóneos para originar una variación prolongada en las neuronas postsinápticas, porque se cierran en cuestión de milisegundos una vez desaparece la sustancia transmisora. Se consigue una excitación o una inhibición neuronal postsináptica a largo plazo al activar un sistema químico de «segundo mensajero» en el interior de esta misma célula, y a continuación será este elemento el que genere el efecto duradero.

- Apertura de canales iónicos específicos a través de la membrana celular postsináptica.
- Activación del monofosfato de adenosina cíclico (AMPc) o del monofosfato de guanosina cíclico(GMPc) en la neurona.
- Activación de una enzima intracelular o más.
- Activación de la transcripción génica.
Excitación:
- Apertura de los canales de sodio para dejar pasar grandes cantidades de cargas eléctricas
positivas hacia el interior de la célula postsináptica. - Depresión de la conducción mediante los canales de cloruro, de potasio o ambos.
- Diversos cambios en el metabolismo interno de la neurona postsináptica para excitar la actividad celular.
Inhibición:
- Apertura de los canales del ion cloruro en la membrana neuronal postsináptica.
- Aumento de la conductancia para los iones potasio fuera de la neurona.
- Activación de las enzimas receptoras.
Los neuropéptidos se sintetizan de otro modo y tienen acciones que normalmente son lentas y en otros aspectos bastante diferentes de las que ejercen los transmisores de molécula pequeña. Estas sustancias no se sintetizan en el citoplasma de los terminales presinápticos. Por el contrario, se forman en los ribosomas del soma neuronal ya como porciones íntegras de grandes moléculas proteicas.
